Aggressivita
Dizionario di Medicina (2010)
aggressività
L’aggressività rappresenta la tendenza di un individuo a esprimere comportamenti ostili verso uno o più membri della propria o di un’altra specie. L’aggressività è molto diffusa in tutto il regno animale, dagli invertebrati all’uomo, dove si manifesta attraverso un repertorio comportamentale ritualizzato. Essa è indotta da particolari stimoli evocatori emessi da uno dei contendenti, che possono essere colorazioni, strutture corporee o sostanze chimiche. L’aggressività ha cause genetiche, ma può essere criticamente modulata dall’esperienza e da fattori ambientali. La regolazione dell’aggressività coinvolge diverse aree del sistema nervoso centrale e di quello endocrino, che interagiscono in modo complesso. L’analisi dei meccanismi alla base dell’espressione dell’aggressività si avvale di studi condotti in laboratorio su animali, che hanno contribuito in maniera fondamentale sia a comprendere l’aggressività umana, sia a identificare strategie volte a ridurre l’aggressività negli animali di allevamento, da compagnia e da laboratorio, con importanti ricadute etiche ed economiche. [➔ comportamento sociale; cure parentali; geni omeotici e cervello; istinti; neurovegetativo, sistema; stress e adattamento] Nella sua accezione più comune l’a. si riferisce alla manifestazione di comportamenti ostili verso qualcuno o qualcosa. Nonostante l’appartenenza di questo termine al linguaggio comune, la definizione del concetto di a. ha costituito oggetto di un intenso dibattito che ha appassionato a lungo un’ampia parte della comunità scientifica e filosofica. Le ragioni di tale dibattito risiedono sia nel carattere ampio e generale del termine, sia nella molteplicità e varietà delle discipline coinvolte in tale dibattito, quali la biologia, l’etologia, la psicologia, la sociologia e la filosofia. Indubbiamente, un buon punto di partenza consiste nel definire l’a. in un contesto biologico: essa si riferisce al comportamento con cui un animale reca danno, reale o anche solo potenziale, a un altro animale. Tale definizione permette di includere gli attacchi falliti e non specifica la natura del danno (che può essere sia fisico sia psicologico). Inoltre, essa prescinde dal tipo di soggetti coinvolti nel comportamento aggressivo: sono infatti da considerarsi forme di a. quelle manifestate verso un animale della stessa o di un’altra specie (rispettivamente, a. intra- o interspecifica), siano esse a scopo predatorio, territoriale, o di difesa della prole.
Caratteristiche del comportamento aggressivo
Il comportamento aggressivo è ampiamente diffuso nel regno animale, presentandosi con una varietà di caratteristiche pari alla diversità esistente tra le specie. Tuttavia esistono caratteristiche del comportamento aggressivo che prescindono dalle differenze specie-specifiche. Innanzi tutto, l’a. è sempre legata alla disponibilità di una determinata risorsa, necessaria ai contendenti. L’a. può essere finalizzata all’ottenimento o alla difesa immediati di cibo o di un partner sessuale specifico, ma anche alla conquista o al mantenimento di un territorio, ossia un’area in cui è molto probabile trovare tali risorse per la sopravvivenza. Numerose teorie sono state utilizzate per spiegare i vantaggi dell’aggressività. Tra queste la più accreditata è la cosiddetta teoria dei giochi, che mette in relazione i costi e i benefici dei comportamenti aggressivi con la frequenza con cui essi si manifestano. Un’altra caratteristica comune dell’a. è che si struttura in un repertorio di segnali comportamentali ritualizzato e stereotipato. Tale repertorio ha alcuni aspetti comuni in tutte le specie animali, quali la presenza di segnali di esibizione espressi a distanza e di azioni che prevedono il contatto diretto, di cui alcune innocue, mirate a scoraggiare il rivale, altre tese al suo ferimento, in alcuni casi fino all’uccisione.
Segnali evocatori
Al pari di altri moduli fissi di attività, anche quelli aggressivi sono evocati da particolari stimoli-segnale emessi dal contendente. Nelle specie che utilizzano principalmente la vista come canale di comunicazione i segnali sono particolari strutture o colorazioni corporee esibite a scopo di minaccia nelle fasi che precedono il combattimento vero e proprio. Gli studi di Nikolaas Tinbergen sui maschi del pesce spinarello hanno mostrato come sia la colorazione rossa del ventre del rivale a scatenare il comportamento aggressivo dell’individuo territoriale. Tale segnale è talmente potente che zimbelli grossolani la cui parte inferiore sia dipinta di rosso funzionano da evocatori più efficaci di sagome non colorate che riproducono fedelmente la forma del rivale. Nelle specie a olfatto sviluppato, tra cui molte specie di mammiferi, i segnali aggressivi sono principalmente di natura chimica: i topi maschi, per es., emettono con l’urina sostanze odorose che scatenano la risposta aggressiva. Insieme ai segnali stimolatori si sono evoluti meccanismi capaci di inibire l’aggressività. Pur nella loro varietà specie-specifica, i segnali di sottomissione nell’ambito dei vertebrati sono spesso simili ai segnali infantili o sessuali. Nell’insieme, la ritualizzazione del comportamento aggressivo ha lo scopo di neutralizzarne la pericolosità evitando, se possibile, esiti letali. Per es., i topi maschi generalmente evitano i morsi al ventre e ai genitali, aree particolarmente vulnerabili all’azione a tenaglia dei loro incisivi. È interessante notare come il contrario accada nei moduli di a. messi in atto dalle femmine per la difesa della prole.
Le cause dell’aggressività: geni e ambiente
Il carattere stereotipato dei moduli di comportamento agonistico depone a favore di una loro coordinazione ereditaria. Tinbergen ha dimostrato come i maschi adulti del pesce spinarello allevati in isolamento sociale fin dalla nascita combattano con la stessa tecnica dei loro conspecifici che hanno avuto precedenti esperienze di lotta. Lo stesso fenomeno è stato riscontrato in altre specie di pesci, rettili, uccelli e mammiferi, topi compresi. In partic., studi condotti sui topi selvatici e sui loro discendenti di laboratorio hanno rivelato la coesistenza, all’interno di una popolazione, di individui che attaccano rapidamente un conspecifico che si introduce nel loro territorio e altri lenti ad attaccare. Tale variabilità intraspecifica può essere spiegata considerando l’organizzazione sociale di questa specie. I topi attraversano fasi di organizzazione stabile e periodi di migrazione. Durante le fasi di stabilità possono alternare un’organizzazione territoriale, in cui i maschi adulti si contendono i territori in cui risiedono diverse femmine mature, a un’organizzazione gerarchica, con dominanti e subordinati. Gli individui rapidi ad attaccare avranno più successo riproduttivo nei periodi di stabilità, mentre quelli lenti saranno favoriti nelle fasi di migrazione. È stato dimostrato come questa divergenza sia di origine genetica, poiché le due diverse linee sono state selezionate nel corso di circa una trentina di generazioni. È interessante notare come i profili ‘lento’ e ‘veloce’, riscontrati anche in altre specie di mammiferi e in alcuni uccelli, non si limitino all’a., ma corrispondano a diverse strategie con cui gli individui affrontano l’ambiente in generale. Tali differenze individuali sono note anche come personalità animali. Nonostante il controllo genetico dell’a. sia stato dimostrato in numerose specie animali, lo sviluppo e l’espressione dell’a. si basano chiaramente su una complessa interazione tra geni e ambiente. L’a. è stata infatti definita come un istinto reattivo che, pur avendo una sua programmazione biologica, si manifesta in risposta agli stimoli che lo evocano ed è rinforzato dall’esperienza. Esperienze compiute durante le diverse fasi della vita di un individuo possono essere cruciali nel modulare la sua aggressività. Alterazioni delle interazioni madre-piccolo, per es., possono modificare potentemente l’espressione dei comportamenti aggressivi e l’acquisizione dei ruoli di dominanza/subordinazione in età adulta in numerose specie di mammiferi. Anche cambiamenti dell’ambiente sociale durante lo sviluppo postnatale o l’età adulta rappresentano importanti fattori di modulazione del comportamento agonistico.
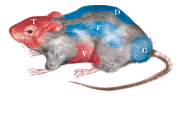
Studi condotti in topi di laboratorio hanno dimostrato, per es., come l’isolamento sociale subito dopo lo svezzamento determini l’apparizione precoce di comportamenti aggressivi. L’isolamento condotto in età adulta determina invece un notevole aumento dei livelli di a. in numerose specie di mammiferi, tanto da essere utilizzato come procedura preparatoria in molti studi di laboratorio. Anche modificazioni più sottili dell’ambiente sociale possono avere un’influenza determinante sull’aggressività. Variare sistematicamente la composizione di una colonia di ratti induce un aumento dei comportamenti aggressivi verso un intruso. Aumentare la densità di una colonia di topi di laboratorio tende a favorire l’organizzazione territoriale rispetto a quella gerarchica, essendo la dominanza eccessivamente difficile da difendere in presenza di numerosi individui. In questa specie, inoltre, combinare la coabitazione di più femmine con un solo maschio induce il manifestarsi di intensa a., un fenomeno che si osserva raramente tra le femmine.
Cervello, ormoni e comportamento aggressivo
Nei vertebrati, alcuni dei substrati neurali principalmente coinvolti nella regolazione del comportamento aggressivo sono stati individuati nelle strutture paleoencefaliche. Tali strutture, le più antiche del sistema nervoso centrale dal punto di vista filogenetico, comprendono la formazione reticolare, che controlla lo stato di attivazione (arousal) dell’organismo, il sistema limbico, il talamo e l’ipotalamo. In partic., studi condotti su mammiferi predatori hanno messo in evidenza come stimolando le aree mediali ipotalamiche si induca una risposta difensiva, quelle dorsali evocano invece risposte di fuga e le laterali il tipico attacco predatorio. Tuttavia l’ipotalamo non è il solo centro nervoso in grado di gestire specifiche risposte aggressive. In diverse specie di mammiferi lesioni dell’aerea ventrale dell’ippocampo impediscono la manifestazione di attacchi predatori che non possono più essere evocati tramite stimolazioni ipotalamiche, mentre lesioni dell’amigdala inibiscono i comportamenti aggressivi di difesa. È importante anche notare come gli effetti di lesioni di alcune aree corticali possano dipendere dallo stato gerarchico dell’individuo. Gli ormoni svolgono un ruolo determinante nella modulazione del comportamento aggressivo. L’influenza del testosterone nel controllo dell’a. intraspecifica tra maschi è ben nota: la castrazione determina infatti una drastica riduzione dell’a., che può essere ripristinata tramite la somministrazione di testosterone in diverse specie di pesci, rettili, uccelli e mammiferi. Tuttavia il livello ematico di testosterone non è sufficiente a determinare l’a. di un individuo. Studi condotti nei primati hanno dimostrato come esperienze di insuccesso in un combattimento possano essere talmente determinanti che nemmeno la somministrazione di testosterone può rendere aggressivo un animale subordinato.
È chiaro dunque come l’interazione tra ormoni e a. sia molto complessa. Innanzi tutto l’azione degli ormoni sul comportamento aggressivo può essere anche indiretta: è stato per es. dimostrato che topi sottoposti a castrazione risultano coinvolti in un numero inferiore di episodi agonistici poiché non producono i segnali chimici che stimolano l’attacco. Inoltre, gli ormoni controllano l’a., ma le esperienze di lotta influenzano a loro volta l’attività ormonale. In partic., poiché i conflitti rappresentano esperienze stressanti, i cosiddetti ormoni dello stress, quali le catecolammine, i glicocorticoidi e l’ormone adrenocorticotropo (ACTH), fanno parte integrante del substrato endocrino del comportamento aggressivo. È stato dimostrato, per es., che nei topi e nei ratti entrambi i partecipanti a un conflitto presentano un incremento nei livelli ematici di glicocorticoidi e ACTH che appare essere più marcato e a lungo termine nei subordinati. L’attivazione dell’asse ipotalamo- ipofisi-surrenali preposta alla secrezione di tali ormoni sembra dunque essere fondamentale per la regolazione dei rapporti di dominanza/subordinazione: la rimozione dell’ipofisi impedisce infatti lo stabilirsi di tali ruoli, mentre la somministrazione di ACTH accentua e accelera il manifestarsi di comportamenti di sottomissione. Infine, altri fattori di natura non ormonale svolgono un ruolo fondamentale nella regolazione dell’aggressività. Il fattore di crescita nervoso (➔ NGF), per es., viene rilasciato dalle ghiandole salivari dei topi maschi esclusivamente in seguito a interazioni agonistiche. L’NGF agisce come agente trofico sia per il sistema nervoso periferico sia per le ghiandole surrenali, la cui morfologia e funzionalità sono modificate dall’esposizione a ripetuti incontri aggressivi. Individui subordinati presentano infatti un’ipertrofia delle ghiandole surrenali accompagnata da livelli ematici di NGF più alti di quelli dei dominanti. L’esperienza agonistica induce anche un aumento dei livelli di NGF nel sistema nervoso centrale dei topi maschi, in partic. nell’ipotalamo, e variazioni nell’espressione del fattore di crescita encefalo-derivato (BDNF, Brain-Derived Neurotrophic Factor) in varie aree cerebrali, per es., nell’ippocampo. I cambiamenti osservati nell’espressione di entrambe le neurotrofine si spiegano come fenomeni di plasticità cerebrale legati all’esperienza agonistica.
Studiare l’aggressività in laboratorio
Le ricerche in laboratorio hanno il vantaggio di consentire la valutazione degli effetti di variazioni genetiche o ambientali in maniera più semplice e standardizzata rispetto agli studi sul campo. In laboratorio è possibile controllare le condizioni ambientali, per es. le dimensioni delle gabbie di mantenimento degli animali, o il numero di individui in ciascun gruppo, così come la luminosità o la temperatura. Inoltre è possibile eseguire studi di selezione genetica o modificare specifici geni. L’a. in laboratorio è stata largamente studiata utilizzando diverse specie di insetti, uccelli, pesci, rettili, crostacei e mammiferi. I topi tuttavia rappresentano sicuramente gli animali più utilizzati, poiché sono facilmente allevabili in condizioni di cattività e utilizzabili per generare linee geneticamente modificate. Inoltre, i topi esprimono un repertorio aggressivo comportamentale ben sviluppato, che si esprime soprattutto nei maschi a partire dall’età di circa un mese. L’a. nei topi maschi è stata studiata principalmente introducendo due animali sconosciuti in una gabbia nuova o un intruso nella gabbia di un residente. Gli incontri in ambiente neutro possono essere ripetuti per diversi giorni; tale procedura è effettivamente necessaria per osservare come si definiscono i rapporti di dominanza/ subordinazione. Il comportamento aggressivo può essere quantificato analizzando la frequenza e la durata dell’espressione delle posture che compongono il repertorio agonistico di questa specie.
Perché studiare il comportamento aggressivo
Gli studi sull’a. hanno importanti applicazioni innanzitutto per la gestione degli animali da compagnia, come sottolineato da alcuni drammatici episodi di cronaca degli ultimi anni. Le ricerche sull’a. hanno anche altre importanti ricadute di tipo etico ed economico: conoscere le strategie per ridurre l’a. negli animali di allevamento o di laboratorio equivale a migliorarne il benessere psicofisico, con chiari vantaggi per la qualità rispettivamente dei prodotti e dei dati sperimentali. Infine, riveste da sempre notevole importanza la comprensione dell’a. umana, sia come espressione di patologie neuropsichiatriche, sia come causa di problemi di natura sociopolitica. Susanna Pietropaolo
Correlati neuroanatomici e biochimici
Sembra che i circuiti nervosi che regolano i comportamenti aggressivi (CA) facciano parte della rete neuronale che media i comportamenti sociali. Tale rete comprende l’area mediale preottica (o AMP, una regione dell’ippocampo che regola l’aggressività tra i maschi, i comportamenti sessuali e le cure genitoriali), il setto laterale (SL), l’ipotalamo anteriore (IA) e ventromediale (IVM), la sostanza grigia periacquiduttale (PAG), l’amigdala mediale (AM) e il nucleo del letto della stria terminale (NLST) e la corteccia orbitofrontale (COF).
L’aggressività negli animali
Nei roditori le informazioni provenienti dal bulbo olfattivo − utilizzate per la ricerca del cibo, l’esplorazione dell’ambiente, il riconoscimento individuale e la comunicazione tra individui della stessa specie − raggiungono l’amigdala mediale (MEA), una struttura deputata a valutare il pericolo e a organizzare il comportamento conseguente. Essa invia proiezioni all’ipotalamo (IA e IVM), il quale regola l’attivazione del sistema nervoso autonomo e quindi le reazioni fisiologiche tipiche del CA e, attraverso l’NLST e l’SL, alla sostanza grigia periacqueduttale del mesencefalo che induce la reazione comportamentale. Sembra che diversi sottonuclei siano più attivi in diversi contesti sociali. Per es., la regione posteroventrale dell’AM e la dorsomediale dell’IVM sono più importanti nel regolare l’aggressività in situazioni di difesa, mentre la regione posterodorsale dell’AM nell’aggressività di tipo offensivo. In generale, lesioni di SL, NLST, IA e AM sopprimono i CA, mentre lesioni della COF aumentano l’aggressività, suggerendo che essa svolga un effetto inibitorio sulla rete neuronale dei comportamenti sociali. La stimolazione elettrica dell’IA (noto anche come area di attacco) aumenta, mentre il blocco dei recettori della vasopressina nei neuroni dell’IA riduce l’aggressività dei maschi della stessa specie. Indagini di tipo immunoistochimico, atte a evidenziare modulazioni dei geni precoci come c-fos, indicano che il prodotto di questo gene aumenta in maniera significativa in SL, NLST, IA e AM in diverse situazioni tra cui: l’aggressività tra i maschi; l’aggressività tra le femmine in relazione alle fasi dell’estro; l’aggressività che insorge nelle femmine durante la gestazione e la lattazione per difendere la prole dai maschi. Nei primati non umani i CA sono indotti da segnali acustici o visivi e, come nel caso dei roditori, l’ipotalamo e l’amigdala svolgono un ruolo chiave. La stimolazione elettrica dell’IVM aumenta i vocalizzi minacciosi e la piloerezione nei maschi delle scimmie marmosette (Callithrix jacchus), mentre lesioni dell’IA e dell’area preottica riducono i vocalizzi emessi alla presenza di un intruso maschio. L’amigdala ha un ruolo più complesso: lesioni dell’AM infatti stimolano o sopprimono i CA tra maschi della stessa specie a seconda che, dopo la lesione, l’animale venga reintrodotto in un ambiente più o meno conflittuale. La COF sarebbe, invece, implicata nell’interpretazione dei ruoli sociali e nelle risposte comportamentali a complesse situazioni sociali: per es., lesioni della COF causano aumenti dell’aggressività nei maschi dominanti e riduzione nei maschi subordinati.
L’aggressività nell’uomo
Nell’uomo, studi di lesione e di visualizzione encefalica (PET, RMN) indicano che i circuiti nervosi che mediano l’aggressività reattiva sono in parte simili a quelli dei primati e dei roditori. Il ruolo critico esercitato dalla COF è stato per la prima volta riconosciuto e messo in evidenza in pazienti con lesioni in questa sede che manifestavano CA. Queste lesioni possono essere il risultato di un trauma, di un tumore o di disturbi metabolici. In genere i pazienti con lesioni del lobo frontale sono più inclini, nelle situazioni di conflitto, a fare uso della intimidazione fisica e verbale. Anche il lobo temporale è implicato nella suscettibilità a violenza e aggressività, come emerso da studi su pazienti con lesioni o tumori in questa sede o con epilessia del lobo temporale. Mediante studi di visualizzazione encefalica, nei pazienti affetti da disturbo antisociale della personalità è stata riscontrata una riduzione significativa del volume della sostanza grigia in sede prefrontale, attribuita a difetti dello sviluppo. L’interpretazione di queste osservazioni deve comunque tener conto e non la conseguenza della malattia.
Correlati biochimici dell’aggressività
Alcune ricerche hanno avanzato l’ipotesi che un incremento dell’attività noradrenergica a livello del locus coeruleus, implicato nel controllo dello stato di vigilanza e nella risposta a stimoli esterni, possa favorire CA diretti contro altri. L’aumento di CA di tipo reattivo è stato ricondotto a una ridotta attività serotoninergica a livello della corteccia prefrontale e dell’amigdala. Bassi livelli di serotonina sono generalmente associati ad alti livelli di impulsività e aggressività. Anche l’equilibrio tra amminoacidi eccitatori e inibitori regola i CA. Un incremento dei recettori per il glutammato sembra facilitare il CA, mentre incrementi del GABA inibiscono i neuroni dopamminergici della via mesocorticolimbica e quindi l’aggressività strumentale. Alcuni studi condotti su pazienti affetti da sindrome da deficit di attenzione e iperattività hanno evidenziato che una reciproca modulazione tra sistema serotoninergico e dopamminergico potrebbe essere cruciale nella patogenesi dei disturbi impulsivi. Studi su modelli animali indicano che la riduzione della serotonina o il blocco dei recettori serotoninergici induce condotte impulsive attraverso la disinibizione delle vie dopamminergiche. Anche diversi ormoni di tipo steroideo influenzano l’aggressività. Il più efficiente mediatore ormonale del CA sia nei maschi sia nelle femmine e il testosterone, mentre il progesterone ha effetti contrari. Tra i roditori l’effetto del testosterone sull’aggressività e variabile e dipende dal profilo genetico.
© Istituto della Enciclopedia Italiana - Riproduzione riservata


