Memoria
Enciclopedia on line
Processo legato alla genesi di una modificazione (traccia mnestica) di un substrato, organico o non, attraverso il quale un determinato effetto persiste e diviene suscettibile di rimanifestarsi nel corso di ulteriori occasioni. In particolare, la funzione psichica, che nell’uomo raggiunge il completo sviluppo, di riprodurre nella mente stati di coscienza passati (immagini, sensazioni, nozioni), di poterli riconoscere come tali e di localizzarli nello spazio e nel tempo.
Psicologia
I processi della memoria
Il funzionamento della m. fa riferimento a un insieme complesso di processi in cui sono coinvolte anche altre funzioni cognitive come l’attenzione, la percezione e tutte quelle abilità che hanno a che fare con l’intelligenza generale. I processi specificamente mnestici sono la codifica delle informazioni, l’immagazzinamento, il consolidamento e il ricordo. Le informazioni entrano nel sistema cognitivo attraverso processi di codifica che possono avvenire sotto il controllo strategico (quindi secondo una pianificazione cosciente dell’apprendimento), oppure in modo automatico, tramite meccanismi inconsci.
I processi di codifica a lungo termine trattano l’informazione che proviene direttamente dai sistemi percettivi o dall’elaborazione della m. di lavoro. In quest’ultima le informazioni sono codificate nel formato fonologico o visuospaziale, a seconda del tipo di stimolo e di sottosistema implicato (v. fig.). Un’attività di ripetizione o ripasso permette il temporaneo mantenimento dell’informazione. Nella m. a lungo termine l’efficienza dei processi di codifica dipende da fattori legati sia alla quantità sia alla qualità del lavoro cognitivo attuato. Gli aspetti quantitativi riguardano: a) la durata della presentazione dello stimolo da apprendere, nel senso che più a lungo si è esposti a un’informazione, maggiore è la capacità di immagazzinarla e ricordarla, anche se il tempo di esposizione necessario varia a seconda del materiale e della modalità percettiva implicata; b) la quantità di ripasso (rehearsal) a breve termine; c) la distribuzione del tempo di apprendimento: sembra più efficace una procedura di apprendimento distribuito piuttosto che intensivo.
Gli aspetti qualitativi si riferiscono invece al tipo di elaborazione impiegata, che non solo può essere intenzionale o incidentale, ma che può essere compiuta a diversi livelli di profondità. L’apprendimento involontario di informazioni, definito incidentale, non sembra meno efficace di quello intenzionale, a meno che per quest’ultimo non intervenga una specifica mediazione dei processi attenzionali con la pianificazione di strategie ad hoc. Ciò che sembra invece cruciale per la codifica è la ‘profondità di elaborazione’, vale a dire la misura in cui il fuoco dell’elaborazione si sposta dagli aspetti percettivi superficiali dell’informazione a quelli concettuali. Quanto più il processo elaborativo si focalizza su aspetti semantici e concettuali, tanto migliore sarà l’apprendimento e quindi il recupero successivo. La codifica semantica di un’informazione è più ricca e più facilmente accessibile anche in ragione del suo legame con altre conoscenze. Infatti, un’altra caratteristica dei processi di codifica è la possibilità di organizzare le informazioni nuove con quelle già presenti in memoria. L’organizzazione delle tracce mnestiche, che può essere suggerita dalla natura delle informazioni da apprendere o può essere imposta dal soggetto, è il fattore che maggiormente incide sulla possibilità di ricordare le informazioni medesime.
I processi di codifica possono utilizzare diversi codici, che vanno intesi come formati in cui la mente immagazzina l’informazione (visivo, acustico, verbale, tattile, semantico ecc.). A conclusione del processo di codifica, l’in;formazione è archiviata nei sistemi cui si è fatto riferimento nella presentazione dell’architettura della m., nella forma di una traccia mnestica. L’espressione traccia mnestica o rappresentazione mnestica è riferita a un costrutto ipotetico, che serve a spiegare le relazioni che si instaurano tra una serie di richieste fatte al sistema cognitivo e le sue risposte. Non è ancora chiara né la localizzazione (in un’area specifica o in varie aree cerebrali) né la natura delle rappresentazioni (a ogni traccia corrispondono uno o pochi neuroni, oppure le tracce sono distribuite a rete nel cervello, costituendo dei circuiti nelle regioni coinvolte nei processi di m.?).
La psicologia cognitiva e le neuroscienze hanno lavorato su questo tema in modo separato; mentre la prima si è soprattutto occupata degli attributi delle rappresentazioni, le seconde si sono focalizzate sui meccanismi cellulari sottostanti la formazione della m. e la localizzazione cerebrale di strutture deputate a specifici tipi di apprendimento (m. di facce, m. di nomi, m. spaziale ecc.). Un tentativo interessante di gettare un ponte tra i due approcci è stato compiuto in ambito psicologico dal connessionismo. All’interno di questo modello, le rappresentazioni sono distribuite in un ampio numero di elementi (neuroni) e la conoscenza inerente a tali rappresentazioni in m. è incorporata non solo negli elementi, ma anche nelle connessioni tra di essi. L’apprendimento implica cambiamenti nella forza di tali connessioni e le rappresentazioni in m. costituiscono un pattern di connessioni dotato di un particolare ‘peso’ all’interno di un più vasto insieme di elementi.
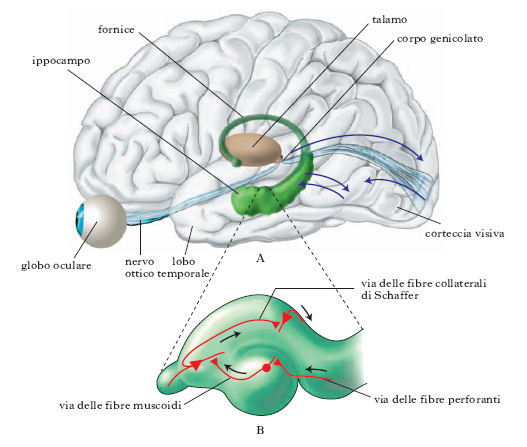
Il destino che le tracce mnestiche subiranno in termini di permanenza e accessibilità al ricordo dipende dal processo di consolidamento, che si presume continui anche dopo che la traccia è stata immagazzinata. Si ritiene che gli eventi che accadono dopo la memorizzazione di un’informazione possano contribuire al proseguimento del consolidamento in m. dei medesimi, o al contrario inibire questo processo creando un’interferenza retroattiva. L’informazione sarebbe dapprima codificata nelle strutture del lobo temporale, principalmente nell’ippocampo (che è danneggiato o asportato nel caso delle amnesie organiche); successivamente, la medesima informazione o parte di essa si stabilizzerebbe in un circuito cerebrale fuori dall’ippocampo, presumibilmente nella neocorteccia. Questo processo di fissazione in altri circuiti avverrebbe in un’estensione di tempo che dipende dalla qualità degli eventi da memorizzare e degli eventi che accadono dopo la prima codifica, a seconda che fungano direttamente o indirettamente da ripasso o che, al contrario, interferiscano per similarità di contenuto.
Ricerche sperimentali
Le prime ricerche
Si fa risalire la storia delle ricerche sperimentali sulla m. ai primi studi dello psicologo H. Ebbinghaus nel 1885. La tecnica messa a punto consisteva nello stabilire il numero di prove o il tempo necessario per l’apprendimento di una serie di liste composte dello stesso numero di ‘sillabe senza significato’. Ciò gli permise in primo luogo di stabilire una curva di acquisizione calcolando il rapporto esistente tra la lunghezza delle liste di sillabe ritenute e il numero di presentazioni necessarie. Egli inoltre utilizzò una tecnica, detta ‘metodo di economia’, per stabilire una curva di ritenzione calcolando il numero di prove necessario al riapprendimento del materiale precedentemente acquisito. Questo metodo di economia nel riapprendimento doveva permettere di riconoscere la fragilità della traccia costituitasi. L’efficacia della ritenzione decresceva infatti rapidamente con il passare delle ore dopo la fine della prova. Alla fase iniziale di rapido declino subentrava una fase di stabilità relativa.
In seguito, un secondo aspetto delle ricerche sulla m. fu chiarito da G.E. Müller e A. Pilzecker (1900), che misero in evidenza gli effetti dei fattori positivi e negativi immediatamente consecutivi all’apprendimento sui processi di ritenzione. Tali fenomeni, che sono stati successivamente descritti con il nome di interferenza, sono stati interpretati dagli psicologi con l’ipotesi della concorrenza tra le informazioni o attraverso l’introduzione del concetto di generalizzazione e di differenziazione.
Tecniche di studio
A seconda del grado di complessità dell’apprendimento considerato, lo studio della m. si avvale di tre gruppi di test: a) le prove di apprendimento motorio basate su test di coordinazione; fra le apparecchiature usate sono i test psicotecnici, simili a quelli per individuare l’abilità alla guida o al tiro a segno; b) le prove di riconoscimento, nelle quali dopo aver presentato figure od oggetti di varia complessità si chiede al soggetto di riconoscere, nel corso di una seconda presentazione distanziata dalla prima da periodi vari di tempo, una serie di figure analoghe; c) le prove di m. verbale, costituite, secondo il principio adottato da Ebbinghaus, dalla riproduzione di un materiale senza significato (serie di numeri, sillabe) o di un testo qualsiasi di cui si chiede di riprodurre il concetto o le parole esatte.
Lo studio dei processi di memorizzazione e dei loro meccanismi nell’animale di laboratorio si avvale delle tecniche di condizionamento classico o pavloviano, del condizionamento strumentale, o dell’uso di molluschi marini, come Aplysia o Hermissenda, o di lumache come Limax maximus.
Ipotesi sulle origini della traccia mnestica
Secondo un’interpretazione fornita da D.O. Hebb (1949), il primo stadio della traccia mnestica corrisponderebbe a modificazioni funzionali neuroniche e si avvarrebbe di circuiti di riverberazione, e cioè della ripetizione di un’attività elettrica attraverso strutture nervose chiuse. A favore di questa teoria sono dati che mostrano come sia possibile rivelare con tecniche elettrofisiologiche dei cicli ricorrenti di onde elettriche a livello del talamo in seguito alla presentazione di stimoli; questi cicli di onde farebbero pensare alla continua circolazione della rappresentazione dello stimolo attraverso una rete chiusa di neuroni. Un singolo stimolo può dar luogo a cicli di onde elettriche che si ripetono per un tempo che può durare 30 minuti e oltre.
Per ciò che riguarda il secondo stadio dell’immagazzinamento della traccia, corrispondente alla m. a lungo termine, sono state prospettate differenti ipotesi secondo cui: a) i ricordi sarebbero basati su modificazioni strutturali e anatomiche del sistema nervoso; b) l’accumulo della m. sarebbe basato, come nel caso della m. transazionale o a lungo termine, su modificazioni chimiche similmente a quanto avviene per la m. genetica; c) in base a un concetto eclettico, la formazione dell’engramma sarebbe costituita da modificazioni chimiche a livello della sinapsi. A favore di un concetto basato su modificazioni strutturali e anatomiche sono da ascrivere i dati filogenetici che mostrano un maggior sviluppo delle diramazioni interneuroniche (sostanza grigia) ai livelli più elevati della scala zoologica e numerosi risultati che dimostrano come l’ambiente e gli stimoli sensoriali esercitino un’azione positiva sulla crescita e lo sviluppo delle cellule e diramazioni nervose. In particolare, le ricerche di M.R. Rosenzweig e collaboratori negli anni 1950 e 1960 hanno posto in evidenza come allevando due gruppi di animali fin dalla nascita in condizioni ambientali completamente differenti, e cioè in un ambiente ‘stimolante’ (per la presenza di piccoli giochi, luci e stimoli auditivi) o ‘impoverito’ (per l’assenza di stimolazioni sensoriali), gli animali del primo gruppo presentano, da adulti, un maggior peso e spessore della corteccia cerebrale, un aumento nella lunghezza e nel numero delle diramazioni neuroniche e infine più elevati livelli dell’enzima acetilcolinesterasi cerebrale. Queste modificazioni sembrano accompagnarsi a un’accresciuta abilità nel risolvere problemi di particolare difficoltà. L’ipotesi secondo la quale gli engrammi che costituiscono la m. corrisponderebbero a un codice prestabilito sotto forma di molecole di RNA è stata formulata da H. Hydén (1962).
Secondo la teoria della consolidazione, la traccia mnestica non viene immagazzinata in modo stabile dopo uno stimolo, ma sarebbe necessario un intervallo di tempo perché la traccia sia ‘consolidata’, cioè immagazzinata in forma durevole. Il modello dualistico della m. (circuiti riverberanti e aspetto neurochimico) è documentato dalle osservazioni che dimostrano la possibilità d’intervenire, dopo la prima fase dell’apprendimento, sui processi che sono alla base della m. a lungo termine. Secondo questo concetto, i fenomeni di amnesia retrograda che sono stati osservati in clinica in seguito a un elettroshock, a un trauma cranico o a una semplice anestesia generale sono interpretati come un blocco di meccanismi di consolidazione. Tali osservazioni cliniche sono state confermate da osservazioni realizzate sull’animale. Anche nell’animale da esperimento si è osservato come, attraverso la somministrazione di alcuni farmaci psicostimolanti come la strictinina o la nicotina, fosse invece possibile innalzare i processi di consolidazione provocando un miglioramento delle prestazioni nelle prove di condizionamento strumentale.
Il concetto della pluralità della m. risulta documentato da ricerche di psicogenetica che hanno dimostrato come vari ceppi puri (inbred) di animali di laboratorio presentino una prevalenza ora dei meccanismi della m. a breve termine ora di quella a lungo termine secondo la natura dell’esercizio. Inoltre, entra in gioco un fattore genetico: alcuni ceppi dimostrano d’imparare meglio un esercizio se le prove sono spaziate nel tempo, mentre altri danno prestazioni migliori se le prove si susseguono rapidamente.
Presupposti anatomici della memoria
Le indagini sul substrato anatomico della m. si sono sviluppate secondo due direttrici: una orientata a individuarlo in limitati distretti cerebrali e una incline a considerare nella m. un intervento diffuso di tutto il cervello, se non dell’intero sistema nervoso. È ovvio che l’evoluzione delle conoscenze in tema d’integrazione funzionale ha interferito sugli indirizzi delle ricerche, confermando o rettificando, a volte smentendo, i dati desunti dall’anatomia clinica e quelli che, in tempi più recenti e con maggior precisione, sono stati forniti dalle osservazioni neurochirurgiche e dalla sperimentazione neurofisiologica.
Lo studio delle localizzazioni della m. umana si è avvalso notevolmente delle osservazioni condotte in pazienti epilettici cui venivano rimosse porzioni del tessuto nervoso allo scopo di bloccare le crisi del male. È tale il caso di un paziente studiato da B. Milner a partire dagli anni 1950, in cui l’operazione distrusse bilateralmente i due terzi anteriori dell’ippocampo e del giro ippocampico, l’uncus e l’amigdala. Guarito dall’epilessia, il paziente mostrò un’assoluta incapacità di formare m. nuove, un danno cioè al processo di consolidazione della memoria. È da notare che l’importanza dell’ippocampo nella memorizzazione era già stata messa in evidenza nel 1898 da V.M. Bechterev, il quale, in un caso di gravi disturbi della m., aveva dimostrato l’esistenza di lesioni bilaterali appunto in questa struttura. Nei soggetti affetti da sindrome di Korsakov si osserva un’amnesia anterograda, relativa cioè agli eventi successivi all’insorgere della malattia. Il danno interessa la regione encefalica, e in particolare i corpi mammillari e il nucleo talamico dorsomediale. Studi di anatomia funzionale hanno messo in evidenza come il circuito trisinaptico, una via unidirezionale che ha inizio a livello della proiezione della corteccia entorinale al giro dentato e che si chiude con proiezioni di ritorno dal campo ippocampico CAl e dai corpi subicolari alla corteccia entorinale, sia rilevante per l’apprendimento e l’imma;gazzinamento dell’informazione.
È interessante notare che osservazioni effettuate su malati di Alzheimer hanno messo in evidenza una cospicua perdita neuronale in corrispondenza dei livelli II e IV della corteccia entorinale con conseguente isolamento dell’ippocampo. I disturbi della memorizzazione che si osservano in tali pazienti sono stati in larga parte attribuiti a questi danni. Il lobo temporale è stato identificato come sede dei processi di memorizzazione anche da osservazioni effettuate da W.G. Penfield (1952) su pazienti epilettici in cui la stimolazione bilaterale di questa struttura determinava un flusso di ricordi organizzati. L.R. Squire e S. Zola-Morgan dagli anni 1980 hanno studiato il caso di un paziente con un’amnesia anterograda molto grave, unita a un’amnesia retrograda estesa a tutto l’arco della sua esistenza. Gli esami radiografici hanno dimostrato che il cervello aveva danni estesi non solo alla regione temporale mediale, ma anche alla corteccia orbitofrontale e al prosencefalo anteriore: il danno riguardava inoltre i lobi temporali anterolaterali. Le osservazioni condotte in questo paziente hanno suggerito l’ipotesi che responsabili del danno alla m. degli eventi passati siano le lesioni combinate del prosencefalo e della corteccia temporale, strutture che costituirebbero una sorta di serbatoio dei ricordi. Per i ricordi recenti, il lobo frontale svolgerebbe una funzione nel loro richiamo. Tale ruolo gli è attribuito sulla base di osservazioni di anatomia clinica e di esperimenti sull’animale (ablazione dei lobi frontali nella scimmia), che dimostrerebbero, in tali condizioni, un’evanescenza dei ricordi recenti. Infine, gli studi condotti da E. Wanington dalla metà degli anni 1970 hanno evidenziato che la porzione inferiore del lobo parietale sinistro sarebbe preposta a un tipo particolare di m., la m. a breve termine uditiva verbale. Disturbi a questa m. insorgono in individui che presentano lesioni, tumori o traumi localizzati in questa regione del cervello.
Neurofisiologia della memoria
Dal punto di vista della neurofisiologia la m. può essere schematicamente divisa in due categorie: la m. dichiarativa e la m. riflessiva.
La m. dichiarativa è quella che può essere descritta verbalmente: consiste nell’abilità a richiamare volutamente e con conoscenza gli avvenimenti trascorsi, le persone conosciute, i concetti appresi. La m. riflessiva è indipendente dalla coscienza o da processi cognitivi come il confronto o la valutazione, si accumula con il susseguirsi di prove successive ed è quella necessaria per apprendere movimenti e azioni ripetitive (per es., andare in bicicletta o guidare la macchina), ma anche regole grammaticali o matematiche. I due tipi di m. possono collaborare nel richiamare il ricordo di azioni passate e la m. riflessiva può aiutare quella dichiarativa.
I circuiti nervosi che sottostanno ai due tipi di m. sono diversi e localizzati in aree diverse del cervello: vi sono lesioni del cervello che compromettono solo un tipo di m. lasciando l’altro tipo integro. Per es., l’amigdala e il cervelletto sono molto importanti per la m. di tipo riflessivo, mentre le strutture diencefaliche, l’ippocampo e il lobo temporale influenzano la m. dichiarativa. Così, individui amnesici per lesioni alle strutture ippocampali possono apprendere e ricordare compiti procedurali complessi, ma, se interrogati, non sono in grado di ricordare di aver lavorato in precedenza su quella procedura. In questi individui i sistemi cognitivi funzionano regolarmente ma, probabilmente, essi non possono accedere ai sistemi di apprendimento che codificano la m. dichiarativa e che hanno sede nelle strutture diencefaliche, ipotalamiche e nel lobo temporale.
Anche le aree associative della corteccia cerebrale sono molto importanti per il mantenimento della m. e la trasformazione della m. recente in m. di lunga durata. La modesta perdita della m. dichiarativa che si ha nell’anziano durante l’invecchiamento fisiologico è probabilmente dovuta a una normale perdita di cellule e a una diminuita efficienza dei circuiti neuronali dell’ippocampo, mentre la severa perdita di m. che si verifica nei pazienti colpiti da malattie neurodegenerative, come la malattia di Alzheimer, è probabilmente dovuta alle gravi alterazioni dei circuiti nervosi e alla perdita consistente di neuroni nella corteccia. La terapia nei due tipi di amnesia deve essere quindi diversa. Mentre la perdita di m. nell’anziano può essere ritardata o diminuita attraverso l’esercizio continuo dei circuiti mnemonici, cui si può associare una terapia farmacologica che aumenti l’efficienza dei circuiti, nel caso di malattie neurodegenerative si prospetta la possibilità di terapie consistenti nel prevenire la morte dei neuroni, per es., somministrando opportuni fattori di crescita, o nel ripristinare le cellule perdute mediante trapianto di cellule neuronali o staminali.
Le basi cellulari e molecolari che possono spiegare l’apprendimento e la m. non sono ancora del tutto note. I due aspetti della m., quella a breve termine e quella a lungo termine, hanno presumibilmente meccanismi diversi; si pensa che la m. a breve termine sia dovuta a modificazione dell’efficienza delle cellule nervose e dei loro meccanismi di comunicazione. Per es., l’esposizione ripetuta a uno stimolo innocuo porta a un fenomeno di abitudine: così un animale, riconoscendo lo stimolo come innocuo, non reagisce più con un riflesso di difesa. Questa abitudine è dovuta a una minore efficienza della trasmissione sinaptica tra i neuroni sensitivi, gli interneuroni e i motoneuroni. Si ha quindi un tipo di apprendimento basato sulla minore efficienza di un circuito. Nel caso invece dell’esposizione ripetuta a uno stimolo nocivo si ha la modificazione opposta, con un’aumentata efficienza della trasmissione sinaptica: l’apprendimento in questo caso è dunque basato sulla maggiore efficienza sinaptica. L’efficienza della comunicazione è data dall’efficienza dei canali ionici, dei recettori e dei sistemi di trasduzione del segnale intercellulare.
Per il passaggio dalla m. a breve termine alla m. a lungo termine si richiede un cambiamento della struttura dei circuiti nervosi con la sintesi di nuove proteine e la creazione di nuove connessioni sinaptiche tra i neuroni. Il passaggio dalla m. a breve termine a quella a lungo termine è graduale e dipende dalla frequenza e dalla intensità con le quali gli stimoli che devono essere ricordati vengono presentati. È anche importante il collegamento fra stimoli diversi. Un fatto, anche se insignificante, si ricorda maggiormente se è avvenuto in collegamento con un evento emozionalmente rilevante. Questi dati comportamentali hanno una loro base cellulare. Si è osservato che nell’ippocampo stimoli ripetuti di una certa intensità o stimoli leggeri associati a stimoli intensi producono modificazioni cellulari nei circuiti nervosi che permangono per lungo tempo. È la cosiddetta long term potentiation (LTP), posseduta da ciascuna delle tre vie sinaptiche principali dell’ippocampo (via delle fibre perforanti, via delle fibre muscoidi, via delle fibre collaterali di Schaffer) e che sembra essere alla base dell’esperienza mnemonica (fig. B). Inoltre si è osservato che inibendo la sintesi proteica si blocca la m. a lungo termine, ma non quella a breve termine.
Un esercizio mnemonico prolungato negli animali aumenta in modo consistente il numero delle connessioni tra i neuroni; al contrario, un periodo di deprivazione sensoriale diminuisce drasticamente le connessioni neuronali. Questi dati indicano che il cervello ha molte possibilità di rimodellamento plastico, sia di breve sia di lunga durata, e che tale fenomeno è influenzabile dall’esercizio, confermando quello che la pedagogia e la psicologia clinico-sperimentale avevano codificato nell’uomo, cioè che il rimedio principale per aumentare la m., anche nell’anziano, non sono i farmaci, ma il continuo esercizio.
Architettura cognitiva della memoria
La ricerca tende ad ammettere l’esistenza di sistemi di m. differenziati che si sarebbero sviluppati per rispondere a scopi e bisogni diversi, anche se, nella maggior parte dei comportamenti, più sistemi sono impegnati contemporaneamente; tuttavia, pur essendo chiaro che esistono diverse forme di m., sostenute da differenti strutture cerebrali, non mancano orientamenti teorici che preferiscono intendere la m. come un insieme unitario e organizzato di funzioni e processi che possono essere attivati in parallelo, ma che non necessitano di un modello a sistemi plurimi. Comunque, la maggior parte degli studi, avvalorata dai risultati di un notevole lavoro sperimentale, ipotizza modelli di struttura della m. che prevedono l’esistenza di più sistemi indipendenti.
Una definizione di sistema appropriata alla m. e che ha potuto imporsi, grazie anche al contributo delle neuroscienze, afferma che un sistema deve poter essere definito non solo in termini di funzioni cognitive e comportamentali, ma anche in termini di strutture e meccanismi cerebrali, tipo di informazione elaborata (percettiva, simbolica, motoria ecc.) e differenze nelle caratteristiche evolutive filogenetiche e ontogenetiche. Inoltre, per essere identificati, i sistemi devono mostrare la possibilità di un funzionamento autonomo attraverso l’individuazione delle cosiddette doppie dissociazioni; di queste si ha prova quando un deficit nelle prestazioni cognitive relative a un sistema non preclude il funzionamento di un altro sistema e viceversa. In considerazione del fatto che molte strutture neuroanatomiche sono al servizio di più sistemi di m. e che più sistemi di m. lavorano in parallelo, lo sforzo scientifico è quello di individuare in che misura i differenti sistemi di m. ipotizzati contribuiscano all’esecuzione delle varie condotte mnestiche.
Nell’ambito della psicologia cognitiva, il primo modello a più componenti a essere formalizzato, a partire dal quale si sono sviluppate altre e più complesse ipotesi di architettura della m., è stato il cosiddetto modello modale (nel senso di vicino alla moda statistica, perché rappresenta la sintesi delle caratteristiche comuni a modelli precedenti). Esso prevede la distinzione di tre ‘magazzini’: a) un registro sensoriale, la cui funzione è quella di trattenere l’informazione percettiva per un tempo brevissimo dopo la sua scomparsa (da 250 ms a 2 s, in relazione alla modalità sensoriale) e di permettere eventuali elaborazioni successive; b) un magazzino a breve termine, a capacità limitata (762 unità di informazione), nel quale l’informazione è trattenuta, per un tempo molto breve (pochi s), in uno stato di accessibilità, per permetterne il passaggio alla m. a lungo termine e/o l’utilizzo per svolgere altri compiti cognitivi; c) un magazzino a lungo termine, che genericamente si assume come luogo dove la conoscenza si organizza in modo permanente, di cui tuttavia non è possibile definire né la capacità, né il tempo di conservazione delle informazioni. A partire da questo primo modello sono state sviluppate formalizzazioni più complesse dei sistemi di m., che sono riuscite a rispondere sempre meglio ai requisiti richiesti dalla definizione stessa di sistema. In tali formalizzazioni, il registro sensoriale non compare più come sistema di m., dacché si preferisce considerare la funzione di trattenimento dell’informazione sensoriale come propria dei processi percettivi. La m. a breve termine è stata riconcettualizzata come m. di lavoro (working memory), laddove la m. a lungo termine è stata suddivisa in quattro sistemi: di rappresentazione percettiva; procedurale; semantico; episodico.
M. di lavoro e m. a lungo termine
La m. di lavoro è intesa come un insieme di meccanismi predisposti per l’elaborazione cognitiva di tipo intenzionale-strategico di input provenienti da fonti percettive e/o da fonti mnestiche a lungo termine. È un sistema predisposto al mantenimento temporaneo e all’elaborazione dell’informazione durante l’esecuzione di compiti cognitivi. Esso consente parallelamente la rappresentazione nella coscienza dei risultati parziali che provengono dai processi cognitivi in atto. Dati clinici e sperimentali hanno suggerito la necessità di considerare questo sistema come a più componenti, secondo un modello tripartito che prevede un ‘esecutivo centrale’, coadiuvato da due sottosistemi di m. temporanea, a capacità limitata, che mantengono l’informazione rilevante per l’elaborazione centrale: il ‘ciclo fonologico’ (phonological loop) e il ‘taccuino visuospaziale’ (visuospatial sketchpad), deputati rispettivamente al mantenimento dell’informazione basata sul linguaggio e di quella visiva e spaziale. I sottosistemi sono tra loro indipendenti, così che, per es., l’immagazzinamento temporaneo dell’informazione linguistica non inibisce né interferisce con l’immagazzinamento contemporaneo dell’informazione visuospaziale. Immagazzinamento ed esecuzione sono meccanismi separati, perciò si è in grado di compiere un’operazione cognitiva anche se uno dei magazzini di m. temporanea è completamente ‘carico’. Questa formalizzazione della m. di lavoro rende ragione del comportamento mnestico di numerosi casi clinici con un deficit alla m. temporanea che, tuttavia, non pregiudica completamente gli apprendimenti né l’intelligenza, cioè la possibilità di operare su informazioni temporaneamente attivate. L’esecutivo centrale ha una pluralità di funzioni: pianificazione dell’azione in funzione di uno scopo; controllo attenzionale sul processo esecutivo; controllo sui processi automatici, una volta che siano stati avviati, con la possibilità di fare proseguire l’azione in modo intenzionale e controllato; recupero delle informazioni necessarie per l’esecuzione di un compito sia dai magazzini temporanei, sia dalla m. a lungo termine. Resta aperto il problema se sia conveniente continuare a considerare l’esecutivo centrale come un sottosistema della m. di lavoro con funzioni multiple, o semplicemente come un agglomerato di processi di controllo che possono agire in parallelo, indipendenti anche se interconnessi. Le localizzazioni neuroanatomiche individuate indicano che le strutture maggiormente interessate dall’attività dell’esecutivo centrale sono i lobi frontali.
La relazione tra la m. di lavoro e i sistemi di m. a lungo termine è definita dallo scambio continuo di informazioni in entrata e in uscita: la m. di lavoro è al servizio dei processi cognitivi che via via si attualizzano nella mente, e deve perciò prelevare informazioni dalla conoscenza specifica o generale che le persone possiedono e integrarle con le nuove informazioni che provengono dall’esterno. L’autonomia della m. di lavoro, e quindi la sua dissociazione possibile rispetto agli altri sistemi a lungo termine, si evidenzia soprattutto nelle amnesie, in cui il funzionamento a breve termine è intatto mentre è deficitario il recupero delle informazioni passate.
Dal punto di vista descrittivo, nei sistemi della m. a lungo termine è possibile operare una distinzione tra i cosiddetti sistemi di m. implicita (sistema di rappresentazione percettiva e sistema procedurale), cioè quei sistemi la cui informazione è immagazzinata e recuperata in modo non consapevole, e quelli di m. esplicita (sistema semantico e sistema episodico), in cui vi è consapevolezza almeno nel momento del ricordo. Il sistema di rappresentazione percettiva riguarda la rappresentazione e conservazione in m. delle caratteristiche percettive degli oggetti (incluse le parole) a livello presemantico, vale a dire a un livello che non implica l’accesso al loro significato. Le sue operazioni sono disconnesse dalla coscienza e i suoi prodotti non forniscono la base per il riconoscimento o la consapevolezza di esperienze precedenti. Si sviluppa precocemente ed è preservato a lungo nel corso della vita. Le regioni cerebrali coinvolte nell’attività di questo sistema sono prevalentemente quelle occipitali.
Il sistema procedurale è invece deputato all’immagazzinamento delle abilità motorie (per es., andare in bicicletta) e delle modalità di esecuzione di attività cognitive e comportamentali (per es., eseguire semplici operazioni di calcolo o andare al ristorante). È caratterizzato da apprendimenti graduali che si automatizzano e mantengono una certa stabilità nel tempo. I gangli della base e il cervelletto sembrano essere le regioni cerebrali maggiormente implicate nel suo funzionamento, mentre non sembrano interessate le strutture ippocampali necessarie per gli altri sistemi a lungo termine.
Il sistema semantico contiene tutta la conoscenza generale del mondo: linguaggio, concetti, algoritmi, informazioni enciclopediche, conoscenza generale di sé. I suoi contenuti rispondono a criteri di verità o falsità e generalmente, per operare, impegna risorse dell’attenzione. Numerosi sono i modelli di rappresentazione delle conoscenze in questo sistema, da quelli classici a rete (concetti, prototipi, reti semantiche) a quelli che fanno riferimento a organizzazioni proposizionali e schematiche. Tutti i modelli, comunque, prevedono un effetto di ‘propagazione dell’attivazione’, vale a dire che ogni qualvolta viene attivata un’informazione si attivano altre conoscenze a essa connesse; questo effetto di diffusione dell’attivazione delle tracce mnestiche è essenziale per spiegare il processo della rievocazione. Le aree cerebrali implicate nel funzionamento del sistema semantico sono i lobi temporali mediali, soprattutto le strutture ippocampali e il lobo frontale sinistro.
Il sistema episodico conserva le conoscenze relative a eventi esperiti personalmente, quindi legati al tempo soggettivo di apprendimento, e possiede pertanto un valore di verità personale. Anch’esso richiede generalmente risorse dell’attenzione ed è suscettibile di trasformazioni, ricostruzioni e perdita di informazioni. Nella teorizzazione di E. Tulving le conoscenze episodiche sono codificate nella forma di ‘engrammi’, che devono essere intesi come insiemi di caratteristiche dell’evento codificato di cui fanno parte anche elementi del contesto ambientale, cognitivo ed emozionale presenti al momento dell’esperienza. Le aree neuroanatomiche interessate sono sempre le regioni temporali mediali, in particolare le strutture ippocampali, il talamo, il giro del cingolo e i lobi frontali.
Diversi dai sistemi di architettura della m. sono i ‘tipi’ o le ‘forme’ della m., che si definiscono in base alla natura del contenuto informazionale e interessano generalmente più sistemi, nel senso che più di un sistema può essere utilizzato per il trattamento di quel tipo di informazione. Ci si riferisce a quella vasta gamma di etichette puramente descrittive che vengono comunemente utilizzate per riferirsi a differenti contenuti di m.: m. autobiografica, m. olfattiva, m. di facce, m. di nomi, m. prospettica (ovvero il ricordo di compiere azioni nel futuro) ecc.
Meccanismi della rievocazione
Una volta immagazzinata, l’informazione può essere rievocata; in questo caso si dice che l’informazione è disponibile e accessibile al ricordo. Può però accadere che l’informazione, pur se codificata, non sia ricordata; in questo caso essa, anche se disponibile, non è accessibile, fenomeno questo che è peraltro frequente nel comportamento quotidiano. La chiave del successo di una condotta di ricordo risiede nell’efficacia di un suggerimento (retrieval cue) che faccia da innesco al meccanismo di recupero della traccia. L’efficacia di un indizio che faccia riaffiorare un ricordo è data dalla sovrapponibilità delle caratteristiche di tale indizio con l’informazione che deve essere rievocata. In altre parole, il suggerimento deve condividere alcune proprietà con l’informazione da recuperare o con il contesto in cui è stata appresa. Tanto più numerose saranno le proprietà condivise, tanto maggiore sarà la probabilità di ricordo. Il ricordo, essendo un processo cognitivo che opera in serie, passo passo, è sempre ricostruttivo, con un margine di fedeltà rispetto alle informazioni apprese che varia a seconda del tipo di conoscenze. Le conoscenze procedurali, essendo automatizzate, vengono recuperate in modo corrispondente all’apprendimento, così come una parte di quelle semantiche, come i concetti, gli algoritmi, il linguaggio.
Il problema dell’infedeltà nella ricostruzione si pone a proposito della m. di eventi: per ricordare un evento recente non ci sono di solito difficoltà nel rievocare i dettagli specifici, ma se l’evento è più lontano nel tempo il ricordo è più povero, e quindi esso dovrà essere in parte ricostruito facendo delle inferenze plausibili basate sulla conoscenza concettuale generale. La relazione tra gli schemi generali di conoscenza codificati nel sistema semantico e il ricordo di eventi specifici codificati nella m. episodica è molto stretta e avviene in una doppia direzione: l’esperienza di un evento arricchisce e modifica lo schema e quest’ultimo ne permette la ricostruzione mediante inferenze. Il risultato è che il medesimo evento verrà ricostruito nel tempo in modo parzialmente diverso e ricodificato insieme ai nuovi dati di contesto, concomitanti al momento della rievocazione. Soprattutto nell’ambito della psicologia della testimonianza, la ricostruttività di questo tipo di m. apre problemi di non poco conto a proposito dell’attendibilità e fedeltà dei resoconti prodotti in sede testimoniale, a prescindere dalla buona fede del teste.
Patologia della memoria
I disturbi della m. derivano essenzialmente da disfunzioni dei gangli della base, solo in parte reversibili, consecutivi a processi patologici (arteriosclerosi e atrofia cerebrale) o a intossicazioni (ossido di carbonio e alcol etilico) o a traumi cranici. Secondo la localizzazione anatomopatologica del disturbo, e quindi a seconda dei correlati neurofisiologici preposti a determinate dimensioni mnestiche, compaiono quadri clinici diversi. In linea generale vale la distinzione fra deterioramento qualitativo (per es., nella sindrome di Korsakov, con o senza confabulazioni) e deterioramento quantitativo (vari tipi di sindromi amnestiche), con la maggiore o minore presenza di ‘schermi’ psicotici (riproduzione parziale dell’avvenimento ricordato).
Disturbi quantitativi
Le amnesie possono dipendere da un grave disturbo della coscienza che impedisce la percezione, e in questo caso non rappresentano un disturbo della m. vero e proprio; altre volte, invece, pur persistendo la capacità di percepire, è alterata la fissazione (amnesia anterograda), oppure la conservazione dei ricordi, com’è il caso delle amnesie retrograde da gravi traumi cranici, o anche la capacità di riproduzione: in quest’ultima evenienza la perdita dei ricordi, che talora può estendersi a tutta la vita precedente, viene reintegrata non di rado spontaneamente, a volte periodicamente, o sotto ipnosi.
Le ipomnesie consistono in un indebolimento semplice e progressivo della m. in cui viene innanzitutto colpita la capacità di fissare nuovi engrammi e si sviluppa una prevalenza dei ricordi più antichi; si osserva, per es., nelle persone anziane. Le ipermnesie vanno distinte in permanenti, che assumono un carattere patologico nella ‘m. prodigiosa’, in genere circoscritta, quale si riscontra non di rado nell’oligofrenia (ipertrofia parziale della m. per la musica, la matematica ecc.), e transitorie o episodiche, accompagnate da un restringimento di coscienza (per es., sonno ipnotico). Aumento della m. di evocazione si osserva in molti stati non patologici, come sotto un’intensa emozione o in grave pericolo di vita.
Disturbi qualitativi
Le dismnesie consistono nella difficoltà di rievocare certe immagini a un momento determinato e si osservano anche nell’uomo normale per nomi propri, melodie ecc. Le illusioni della m. (o allomnesie) si hanno quando i ricordi sono falsati o inadeguati, specie riguardo alla localizzazione nel tempo e nello spazio. Le pseudomnesie o paramnesie o allucinazioni della m. sono caratterizzate dalla neoformazione di ricordi: vi rientrano i falsi riconoscimenti e i falsi ricordi. I primi comprendono il fenomeno del già vissuto o del già visto e la cosiddetta m. reduplicante, caratterizzata dalla sensazione di avere già attraversato una determinata esperienza; i secondi consistono nell’attribuzione del significato di ricordo a quello che non è altro che un prodotto della fantasia.
Biologia
M. cellulare
Tipo di regolazione dei geni di una cellula che non si modifica nelle generazioni cellulari successive; ne è un esempio la metilazione del DNA. M. immunologica Caratteristica funzionale del sistema immunitario nell’ambito della risposta immunitaria specifica (➔ immunità) per cui i linfociti conservano, dopo il contatto con l’antigene, la capacità di riconoscimento dello stesso anche a distanza di tempo; tale proprietà costituisce un forte strumento di difesa organica, e viene utilizzata nelle vaccinazioni.
Filosofia
Nella filosofia antica la nozione di m. si connette con il problema della possibilità di un sapere inconscio cui essa dà luogo. Alla sua soluzione sono legate sia la concezione platonica sia quella aristotelica.
In Platone la m. (μνήμη) è una sorta di serbatoio delle conoscenze, in cui l’anima accoglie e ritiene le impressioni; sulla differenza tra questa m. inconsapevole e la reminiscenza consapevole (ἀνάμνεσις), attivata in occasione dei richiami associativi della conoscenza sensibile, Platone imposta poi la sua dottrina sulla conoscenza delle idee come reminiscenza di un sapere che l’anima avrebbe acquisito nella sua precedente esistenza.
Pur senza aderire alla dottrina gnoseologica platonica dell’anamnesi, Aristotele riprendeva la distinzione di Platone su basi psicofisiologiche nel De memoria et reminiscentia, dove la m. è concepita come una funzione che conserva e fissa in immagini i dati del senso, e la reminiscenza come la ricerca attiva di quelle immagini, che si attua secondo un ‘movimento’, così come un movimento aveva dato luogo, tramite la sensazione, all’impronta nell’anima. La concezione della m. come serbatoio di rappresentazioni di derivazione sensoriale è posta in discussione da Plotino, che fa della m. un’attività di esclusiva pertinenza dell’anima, indipendente dalle immagini e dalle rappresentazioni connesse alla sensibilità. In s. Agostino, in cui accenti platonici si mescolano a influenze plotiniane, torna una concezione della m. come luogo in cui si conservano le conoscenze, e questa immagine rimarrà praticamente immutata nel pensiero medievale.
È solo in epoca moderna che essa comincia a essere posta in discussione, soprattutto da parte di J. Locke, per il quale la m. consiste nella capacità di far rivivere idee già avute, capacità che si accompagna alla percezione di averle avute in precedenza: si tratterebbe quindi di una nuova percezione, non già della percezione originaria conservata. Se Locke riduceva la m. a un potere di rivivere percezioni già avute, a G.W. Leibniz e T. Reid questo potere sembrava inconcepibile senza presupporre qualcosa che si conserva in seguito alla percezione. E lo stesso I. Kant non faceva che riprendere la concezione aristotelica parlando della m. come della capacità di riprodurre volontariamente rappresentazioni avute in precedenza.
Nel pensiero contemporaneo il concetto di m. come luogo di conservazione, benché non di tipo fisico, è presente in H.-L. Bergson, che parla di una m. ‘pura’ come luogo psichico e spirituale in cui si conserva in uno stato virtuale l’intera vita vissuta. Schiettamente psicologistica, e anch’essa non estranea al modello del deposito, è la concezione esposta da B. Russell in The analysis of mind, dove la m. è definita come «una parte della nostra conoscenza del passato» e il ricordo come un’immagine mentale di derivazione sensoriale accompagnata da un «sentimento di credenza» relativo a episodi passati. In una prospettiva antimentalistica e antipsicologistica che accantona l’immagine della m. come ritenzione di rappresentazioni si collocano poi le concezioni di G. Ryle e L. Wittgenstein: per il primo ricordare non implica una peculiare immagine mentale di ciò che viene ricordato, ma una capacità di fedele riproduzione di ciò che si è visto, udito ecc.; nella concezione di Wittgenstein le nozioni di m. e di ricordo non rinviano a un insieme di rappresentazioni mentali private e soggettive, ma alla capacità socialmente acquisita di usare certe espressioni linguistiche a fini comunicativi.
Informatica
In un sistema elettronico, dispositivo atto a conservare informazioni e a renderle accessibili quando richiesto. Le m. elettroniche sono elementi essenziali nei calcolatori e in molti apparati per telecomunicazioni. Esistono diverse dimensioni per classificare i vari dispositivi di m.; in funzione della combinazione delle possibili scelte lungo le varie dimensioni, si hanno m. dalle caratteristiche e dalle prestazioni molto diverse.
Le tecnologie utilizzate per realizzare dispositivi di m. sono sostanzialmente tre, che portano a una prima classificazione delle m. in elettroniche, magnetiche e ottiche. Nelle prime si sfruttano le caratteristiche proprie dei componenti elettronici che le costituiscono (quali, per es., la carica di un condensatore) per memorizzare l’informazione. Nelle seconde l’informazione è memorizzata mediante opportuna magnetizzazione di supporti appositi, come avviene, per es., negli hard disk. Nelle m. ottiche, infine, vengono memorizzate le informazioni modificando la superficie riflettente delle stesse; esempio tipico sono i comuni dischi ottici, CD e DVD.
M. ROM e PROM
Una delle caratteristiche peculiari delle m. è la modalità con la quale le informazioni possono essere scritte (memorizzate) su di esse; si distinguono m. sulle quali si può scrivere (teoricamente) un numero infinito di volte e m. che possono essere scritte una sola volta, dette ROM (sigla di read only memory). Tipicamente le informazioni presenti su una ROM sono scritte (per tali m. si usa spesso il termine programmate) durante il processo produttivo o comunque prima di arrivare, integrate in qualche dispositivo, all’utente finale; si utilizzano per memorizzare informazioni che non richiedono modifiche per il corretto funzionamento del dispositivo nel quale sono impiegate; per es., si utilizzano delle ROM per memorizzare il firmware o il BIOS di un calcolatore.
Alternativamente, esistono m. ROM che possono essere programmate dall’utente finale una sola volta, dette per tale caratteristica PROM (sigla di programmable ROM).
Esistono altri tipi di m. che sono considerate appartenenti alla categoria delle ROM, pur non rientrandovi strettamente, perché il processo di riscrittura (cancellazione e programmazione) richiede l’uso di strumenti e metodologie apposite; esempi di tali m. sono le EPROM (sigla di erasable programmable ROM), programmabili dall’utente e cancellabili mediante esposizione a raggi ultravioletti, e le EEPROM (electrically erasable programmable ROM), programmabili e cancellabili dall’utente mediante sollecitazione con opportune tensioni.
Volatilità
Le m. sono caratterizzate anche dalla volatilità. L’informazione memorizzata in una m. non volatile è mantenuta per un tempo indefinito senza dispendio di energia; al contrario, una m. si dice volatile se le informazioni in essa contenute si perdono qualora la m. non sia opportunamente alimentata elettricamente o, comunque, vengano meno certe condizioni di funzionamento; la procedura di riscrittura delle informazioni a intervalli regolari prende il nome di refresh.
Modalità di accesso e RAM
Altra caratteristica fondamentale delle m. è la modalità di accesso: si distinguono m. ad accesso sequenziale, che devono essere necessariamente scandite fino al raggiungimento dell’indirizzo relativo all’informazione ricercata da parte del dispositivo di lettura/scrittura (esempio tipico sono i nastri magnetici), e ad accesso casuale dette brevemente RAM (sigla di random access memory), che possono essere lette e scritte a qualunque indirizzo. Le RAM possono essere statiche (non volatili) e dinamiche (volatili); le prime sono dette semplicemente SRAM (sigla di static RAM); le seconde sono ulteriormente divisibili in due categorie, asincrone e sincrone: in quelle asincrone, dette DRAM (sigla di dynamic RAM), le operazioni di lettura e scrittura sono comandate direttamente dai segnali in ingresso mentre in quelle sincrone, dette SDRAM (sigla di synchronous dynamic RAM), tali operazioni sono sincronizzate con il segnale di clock; le DDR (sigla di double data rate) SDRAM sono SDRAM che permettono lettura e scrittura sia sul fronte di salita sia sul fronte di discesa del ciclo di clock.
Capacità
Dal punto di vista funzionale, le m. si distinguono a seconda della loro capacità e del loro tempo di accesso. La capacità di una m. è data dalla quantità di informazione che è possibile memorizzare, misurata in bit. Tipicamente le celle di m. sono costituite da insiemi di 8 bit, corrispondenti a 1 byte, e in genere l’intero organo di m. è caratterizzato da una capacità pari a un numero di byte dato da una potenza di due. Per es., una m. da 1 MB non contiene esattamente 1 milione di byte, ma ne contiene 220, e cioè 1.048.576.
Per individuare una cella di m. si utilizza un sistema di indirizzamento: ogni cella ha un suo indirizzo espresso da un numero binario corrispondente alla sua posizione. Nell’organo di m. o in organi ausiliari è presente un apposito decodificatore per effettuare il corretto indirizzamento.
Dispositivi di memoria
Capacità e tempi di accesso sono due dei fattori che influenzano maggiormente il costo di produzione di una m. e, conseguentemente, il suo prezzo. È anche, e soprattutto, per ottenere il giusto compromesso tra costi e prestazioni che in un calcolatore si è consolidato l’uso di una struttura gerarchica dei dispositivi di memoria. Una versione semplificata di tale gerarchia è costituita da 4 livelli: a) registri; b) cache; c) memoria centrale; d) dispositivi di archiviazione di massa. I registri sono interni alla CPU e offrono le migliori prestazioni al maggior costo unitario. La cache, anch’essa interna alla CPU, tipicamente realizzata con memorie SRAM, serve per minimizzare il numero di accessi alla memoria centrale: in essa si memorizzano dinamicamente le porzioni (pagine) di memoria centrale usate più frequentemente e recentemente; per migliorare ulteriormente le prestazioni, alla cache interna di primo livello si affianca una cache di secondo livello meno performante ma più grande. La memoria centrale, generalmente detta RAM e realizzata con memorie DDR SDRAM, è direttamente accessibile dal processore e in essa devono essere memorizzate le pagine dei programmi che il calcolatore deve eseguire. I dispositivi di archiviazione di massa, come lascia intuire il nome, sono utilizzati per archiviare in maniera stabile grandi quantità di dati; i più comuni sono gli hard disk ma sono utilizzati anche dispositivi di archiviazione a nastro magnetico.
Dimensioni
Orientativamente, gli ordini di grandezza delle dimensioni tipiche delle varie memorie indicate per un PC di fascia medio-alta sono: 32 o 64 bit per un singolo registro, in numero variabile da processore a processore; un paio di centinaia di kilobyte per la cache di primo livello (128 kB - 256 kB); alcuni megabyte per la cache di secondo livello (1 MB - 4 MB); qualche gigabyte per la memoria principale (512 MB - 4 GB); diverse centinaia di gigabyte per i dispositivi di archiviazione di massa (500 GB - 1000 GB).
Letteratura
Nel linguaggio della critica letteraria si designa con le espressioni letteratura della m. e simili quella letteratura per lo più in forma narrativa che in Francia e altrove appare rivolta (sull’esempio di M. Proust e per influsso freudiano) all’evocazione o ricerca del ‘tempo perduto’. Una letteratura analitica, introspettiva, autobiografica di un’autobiografia più fantastica o vagheggiata che realmente vissuta, e più di sensazioni che di pensiero, piena di richiami alla natura, al paesaggio, alle cose, mirante a dar volto ed espressione all’inconscio.
Promossa soprattutto dalla rivista Solaria, una narrativa della m. fiorì anche in Italia, specialmente negli anni tra le due guerre, ed ebbe i suoi principali rappresentanti in A. Bonsanti e nelle scrittrici G. Manzini e A. Banti.
Storia
Giorno della m.
Il 27 gennaio, data dell’abbattimento (nell’anno 1945) dei cancelli di Auschwitz, riconosciuto dalla Repubblica Italiana con l. 211/20 luglio 2000 come giorno destinato a ricordare la Shoah (sterminio del popolo ebraico), le leggi razziali, la persecuzione italiana dei cittadini ebrei, gli Italiani che hanno subito la deportazione, la prigionia, la morte, nonché coloro che, anche in campi e schieramenti diversi, si sono opposti al progetto di sterminio e, a rischio della propria vita, hanno salvato altre vite e offerto protezione ai perseguitati.


