Origini dell'uomo
Enciclopedia della Scienza e della Tecnica (2007)
Origini dell'uomo
Rispetto alle altre discipline scientifiche, l'antropologia presenta una rara anomalia, ovvero la completa circolarità tra osservato e osservatore. L'uomo è al tempo stesso oggetto di studio e soggetto studiante, in una peculiare variante del concetto di indeterminazione in termini epistemologici e del conflitto di interesse in termini storici. L'antropologia è definita come 'storia naturale dell'uomo' o, con un linguaggio più attuale, 'storia naturale del genere umano'. Il problema di un inquadramento naturale della nostra specie si perde nella notte dei tempi, con referenti d'eccezione quali Aristotele o Galeno. Ma il periodo che prepara storicamente l'approccio naturalistico contemporaneo in tal senso è il XVIII sec., dove troviamo scritti, quali la Historie naturelle, générale et particulière di Georges-Louis Leclerc de Buffon, che identificano la natura umana come parte di una natura condivisa con il resto degli organismi viventi. Se è condiviso il fenomeno, i suoi elementi sono comparabili, cioè confrontabili.
L'antropologia si configura come disciplina dall'inizio del secolo successivo, grazie al contributo di personaggi come Johann Friedrich Blumenbach, naturalista tedesco che introduce la misurazione come strumento di confronto per lo studio della variabilità dei gruppi umani. Sono i primi passi di quella che, per necessità di distinzione pragmatica dalla controparte culturale, verrà chiamata 'antropologia fisica' (denominazione sostituita negli ultimi decenni da quella di 'antropologia biologica'). Nella seconda metà del XIX sec. Charles Darwin e i suoi contemporanei introducono le basi delle attuali impostazioni evoluzionistiche, e Thomas Henry Huxley dichiara senza mezzi termini che l'attuale specie umana va considerata integralmente come risultato di un processo di selezione ed evoluzione di forme precedenti. La prima cattedra italiana di Antropologia viene istituita nel 1860 dall'Università di Pavia; a Torino nel 1864 le idee evoluzionisitiche fanno il loro ingresso ufficiale nel nostro Paese con una conferenza di Filippo de Filippi intitolata L'uomo e le scimmie. Nel 1856 viene recuperato vicino Düsseldorf, in Germania, il primo reperto attribuibile a Homo neanderthalensis; nel 1891 l'olandese Eugène Dubois trova nell'isola di Giava il primo esemplare di Homo erectus (o Pitecantropo); nel 1924 Raimond Dart rinviene in Africa meridionale il primo fossile di Australopithecus. Queste date e questi reperti vengono in genere assunti storicamente e simbolicamente come gli esordi più o meno ufficiali della paleoantropologia e degli studi moderni sull'evoluzione dell'uomo.
Attualmente lo studio dell'evoluzione del genere umano rappresenta, in termini sia teorici sia metodologici, un settore estremamente diversificato ed eterogeneo. Lo studio della morfologia e dell'anatomia si avvale delle tecniche digitali, con applicazioni che includono la tomografia computerizzata, l'elaborazione virtuale di immagini, la statistica multivariata e la modellizzazione geometrica delle componenti strutturali. Molti settori della biologia si sono potenziati o aggiunti alle discipline evoluzionistiche, come la paleoneurologia per lo studio delle strutture cerebrali o la paleoistologia per quello delle tracce cellulari fossilizzate. Le tecniche di datazione diretta e indiretta seguono gli evidenti aggiornamenti delle discipline fisiche e chimiche. L'archeologia sottopone a verifiche sperimentali e a modelli quantitativi le sue informazioni storiche. La tecniche genetiche e molecolari riescono a estrarre il DNA rimontando nel tempo fino a circa 40.000 anni fa.
L'evoluzione dei Primati
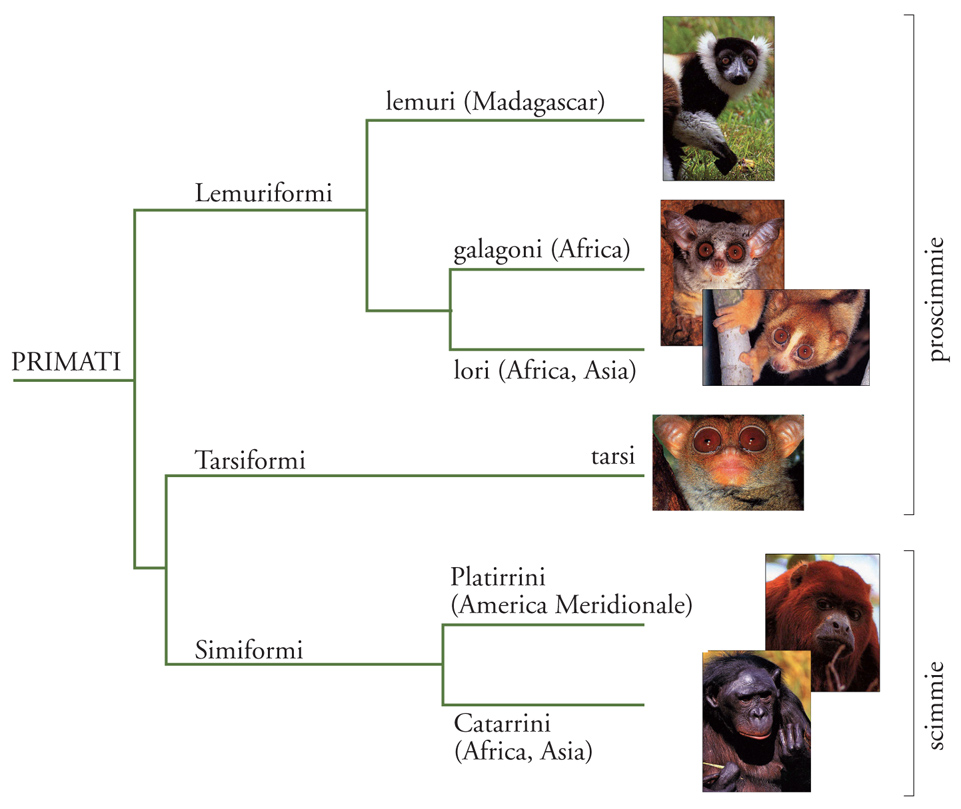
Seguendo l'insostituibile principio di comparazione, fondamento delle scienze evoluzionistiche, per comprendere la storia naturale del genere umano dobbiamo inquadrarlo nel suo gruppo naturale, i Primati. Secondo i raggruppamenti propri delle discipline zoologiche (tassonomia), Primates è uno degli ordini all'interno della classe dei Mammiferi. I Primati contano attualmente più di 350 specie viventi descritte, distribuite soprattutto nelle aree intertropicali. Questo ordine è diviso in tre gruppi principali: Simiformi, Lemuriformi e Tarsiformi (fig. 2). I Simiformi (conosciuti anche con il termine 'Antropoidei') rappresentano le scimmie in senso stretto, con un gruppo afroasiatico (Catarrini) e un gruppo originario dell'America Centrale e Meridionale (Platirrini); i Lemuriformi includono i lemuri (Madagascar), i galagoni (Africa) e i lori (Africa e Asia); i Tarsiformi includono solo i tarsi, con un numero esiguo di specie notturne localizzate nel Sud-Est asiatico. Lemuriformi e Tarsiformi vengono spesso indicati con il termine generico di proscimmie, anche se la loro storia evolutiva è stata relativamente indipendente.
L'evoluzione dei Primati inizia circa 70-60 milioni di anni (m.a.) fa tra l'Europa e l'America Settentrionale, all'epoca non ancora separate geograficamente. Purtroppo il record fossile paleoprimatologico è molto scarso, a causa delle dimensioni corporee spesso modeste delle specie e soprattutto degli habitat comunemente utilizzati da questo gruppo (come quello di foresta), nei quali il processo di fossilizzazione è improbabile e piuttosto raro. Secondo stime orientative potrebbero a oggi essere conosciute solo dall'1 al 3% delle specie di Primati realmente vissute, il che rende il lavoro del paleoprimatologo alquanto affannoso. L'origine dei Primati è da ricondursi a forme di 'protoinsettivori', ovvero piccoli Mammiferi notturni simili come modello biologico agli attuali toporagni o alle tupaie del Sud-Est asiatico. I Plesiadapiformi sono una prima forma zoologica probabilmente riconducibile al modello dei Primati, che popolò l'Europa e l'America Settentrionale nel Paleocene (intorno ai 65-55 m.a.). In termini di aspetto generale, avevano una morfologia e uno stile di vita molto simili ad alcuni Roditori moderni. Nel periodo successivo (Eocene, 55-35 m.a.) queste forme si estinguono e sono sostituite da un gruppo che invece presenta caratteristiche più proprie di un primate (e in particolare delle proscimmie), gli Adapiformi. Nel periodo seguente (Oligocene, 35-25 m.a.) troviamo invece in Africa gruppi di specie con molti caratteri affini alle moderne scimmie platirrine e catarrine.
Definire cosa siano i Primati è compito arduo, per una ragione piuttosto paradossale: essi costituiscono un gruppo primitivo. Ciò equivale a dire che, a fronte di alcune piccole seppure rilevanti specializzazioni, una larga parte della loro struttura biologica è comune ai Mammiferi in generale e alle loro forme originarie, non permettendone una distinzione univoca. Almeno due gruppi di processi sono però utili allo scopo. Il primo caratterizza i Primati per una tendenza alla perdita di potenza delle capacità olfattive, fondamentali per i Mammiferi, in favore di un potenziamento delle capacità visive. Le conseguenze si osservano sia nella morfologia del cranio (accorciamento del blocco facciale e frontalizzazione delle orbite) sia nel sistema nervoso centrale (riduzione dei lobi olfattivi, potenziamento delle aree per l'integrazione visiva). Si perde insomma un mondo fatto di odori a vantaggio di un mondo di forme, colori e distanze.
Il secondo processo riguarda una tendenza all'encefalizzazione, ovvero a un aumento del volume cerebrale in rapporto alle dimensioni dell'individuo. Il cervello è un organo molto dispendioso, ma questo aumento nei Primati avviene senza un associato incremento del sistema energetico (metabolismo). Di conseguenza, se la percentuale maggiore dell'energia va al cervello, il resto del corpo (in particolare il sistema digerente) ne subirà una certa sottrazione. Questa restrizione calorica ha probabilmente prodotto un rallentamento dei processi metabolici nei Primati proporzionalmente alla loro encefalizzazione, con conseguente dilatazione dei tempi vitali (prolungamento della gestazione, dello sviluppo in genere, e della vita media). Le ipotesi evolutive che cercano di interpretare gli adattamenti dei Primati sono differenti, e includono la vita e la locomozione arboricola, la predazione di invertebrati o la dipendenza da fonti nutritive vegetali (foglie, frutti, ecc.). Certamente il modello dei Primati non è il risultato di un solo fattore di selezione: occorre considerare scenari più articolati che tengano conto sia di tutte queste variabili sia di altre fondamentali, quali quelle correlate alla loro peculiare e complessa socialità.
Grandi scimmie
Nel gruppo delle scimmie afroasiatiche (Catarrini) possiamo distinguere due superfamiglie: Cercopithecoidea e Hominoidea. La prima include colobi, macachi, cercopitechi, babbuini e forme affini; alla seconda appartengono le cosiddette 'scimmie antropomorfe' e le grandi scimmie, comprensive della specie umana. L'origine delle scimmie antropomorfe risale al Miocene (23÷5 m.a.), periodo nel quale troviamo i resti fossili di un gruppo chiamato Driopitecine. In questo periodo e nel successivo (Pliocene) esse sono distribuite in Africa, Asia ed Europa, con almeno sette generi attualmente riconosciuti. Una delle caratteristiche principali del gruppo è la tendenza alla locomozione e alla postura verticali. A eccezione di alcune proscimmie e di molte forme estinte che presentano una locomozione saltatoria, molti Primati hanno infatti sviluppato una locomozione quadrupede (arborea o terrestre) nella quale l'asse del corpo è orizzontale (locomozione pronograda). Al contrario, dal Miocene osserviamo che queste specie presentano adattamenti alla brachiazione o alla sospensione, con relativa postura verticale (locomozione ortograda): allungamento degli arti anteriori rispetto ai posteriori, curvatura delle falangi, allargamento del torace e attaccamento posteriore delle scapole, allargamento del bacino, verticalizzazione nel posizionamento della testa.
Attualmente esistono cinque generi di scimmie antropomorfe: scimpanzé, gorilla, oranghi, gibboni e, ovviamente, l'uomo. Scimpanzé (genere Pan; due specie) e gorilla (genere Gorilla; una specie) vivono nell'Africa intertropicale, e hanno parzialmente perduto le specializzazioni sospensorie a favore di una postura inclinata (clinograda), che li vede muoversi frequentemente a terra poggiando sulle seconde falangi della mano. L'orango (genere Pongo; probabilmente una sola specie) vive nelle isole del Borneo e di Sumatra e presenta tutte le caratteristiche tipiche della locomozione sospensoria. I gibboni (genere Hylobates; forse fino a quattordici specie differenti) vivono nel Sud-Est asiatico e rappresentano l'estremizzazione degli adattamenti brachiatori e sospensori. Il genere umano (genere Homo; una sola specie attuale) ha trasformato la sospensione in bipedismo, approfittando degli adattamenti già disponibili alla postura verticale della brachiazione ma potenziando la funzione strutturale degli arti inferiori.
I rapporti evolutivi tra queste scimmie antropomorfe viventi non sono stati ancora del tutto interpretati, soprattutto per quanto riguarda le specie africane. La separazione tra quelle di dimensioni minori (gibboni) e le altre è avvenuta forse intorno a 25-20 m.a. fa e le grandi scimmie asiatiche (oranghi) si sono separate da quelle africane intorno a 20-15 m.a. dal presente. Le dinamiche interne al gruppo africano (Homo, Gorilla, Pan) sono ancora da chiarire. Un'interpretazione biogeografica vede gorilla e scimpanzé separarsi a ovest dai primi Ominidi per l'azione della Rift Valley e delle conseguenze climatiche. Ma le evidenze biologiche indicano invece situazioni più complesse, con dati sia morfologici sia genetici che possono supportare una maggiore prossimità filogenetica tra uomo e scimpanzé o tra scimpanzé e gorilla, a seconda delle variabili considerate (i caratteri usati per i confronti) o delle assunzioni a priori su cui si sviluppano le analisi (le ipotesi di partenza su cui lo studio viene condotto). Anche la rinomata corrispondenza tra il DNA del genere umano e quello dello scimpanzé viene continuamente ridiscussa in termini quantitativi e non fornisce comunque indicazioni sul processo evolutivo, ma solo una stima di similitudine. Non è da escludere comunque che, come in altri casi noti di separazione evolutiva tra gruppi zoologici, il processo sia stato non necessariamente dicotomico (una specie divisa in due specie) ma multiplo, e che i tre generi africani rappresentino quindi una effettiva tricotomia residua di una frammentazione biogeografica avvenuta in un periodo compreso tra 10 e 5 m.a.
Australopitechi

Il termine Ominidi (dal nome della famiglia Hominidae) è utilizzato a volte per indicare quel gruppo naturale che oltre al genere umano include anche lo scimpanzé e il gorilla, volendo sottolineare la coerenza evolutiva di questa triade zoologica. In alternativa, il termine si può limitare a includere il solo genere umano e le forme estinte associate alla sua evoluzione, per dare valore alle peculiarità di questo gruppo rispetto al modello zoologico delle scimmie antropomorfe africane, più simile a quello di altri generi come l'orango. Il primo approccio è più indicato per sottolineare il valore di parentela tra queste specie viventi, mentre il secondo è più corretto quando si voglia dare alla nomenclatura un valore di comparazione biologica. Seguendo questo secondo approccio, la famiglia Hominidae è divisa in due sottofamiglie, Australopithecinae e Homininae. La prima rappresenta le forme estinte non incluse nel genere Homo, mentre la seconda raggruppa tutte le forme umane, estinte e attuali (fig. 3).
Le Australopithecinae si sono evolute e diversificate integralmente in Africa. Per il periodo compreso tra 7 e 5 m.a. dal presente le evidenze fossili sono rare e molto frammentarie e la nostra conoscenza è limitata ad almeno due o tre specie probabili ma non sufficientemente conosciute. È successivamente ai 5 m.a. che il record fossile offre interpretazioni più valide. Se infatti all'inizio del secolo scorso si pensava agli Australopitechi come ad 'anelli mancanti' di un'evoluzione umana graduale e lineare, oggi l'ipotesi comune è quella di un gruppo molto diversificato in termini biogeografici, all'interno del quale solo alcune forme possono essere ritenute ancestrali al successivo genere umano. Sono note almeno 7-8 specie distinte, divise in due gruppi: forme gracili (genere Australopithecus) e forme robuste (genere Paranthropus). Il record fossile è limitato all'Africa orientale e meridionale, con alcuni dati frammentari e non pienamente confermati per i territori a ovest della Rift Valley.
Alcune indicazioni suggeriscono potenzialità bipedi (orme fossili su suoli vulcanici, l'anatomia del femore, la posizione del capo sopra le vertebre cervicali), mentre altre fanno ipotizzare una locomozione ortograda non specializzata. Tali specie avevano infatti arti superiori molto lunghi e falangi ricurve, polsi adatti a sostenere il peso del corpo a terra e strutture dell'orecchio interno (preposte al senso dell'equilibrio) simili a quelle delle attuali scimmie antropomorfe africane. Sono state probabilmente questa non specializzazione e questa generalizzata struttura ortograda a favorire in seguito un pronto adattamento alla locomozione bipede. Gli Australopitechi frequentavano terreni aperti o boscosi, ma probabilmente vivevano in quelle zone di transizione tra habitat con maggiore copertura vegetazionale e aree di savana, ottimizzando le risorse di cibo e protezione di entrambi gli ambienti. Le forme robuste sono definite tali per il loro massiccio apparato masticatorio (molari, zigomi, creste per gli attacchi muscolari sul cranio), probabilmente associato a una dieta caratterizzata da cibi coriacei, più tipici delle aree aperte. In genere, comunque, tutti gli Australopitechi dovevano presentare una dieta appropriativa e onnivora, che includeva componenti vegetali e animali.
La capacità cranica era lievemente superiore a quella delle scimmie antropomorfe attuali, con valori di 400÷600 cm3. Non sono attualmente note per queste forme evidenze culturali come industrie litiche, o le particolari organizzazioni sociali ipotizzate per le specie umane. Forme gracili e forme robuste, a meno di un'origine comune tra 4 e 3 m.a., hanno poi seguito una diversificazione filogenetica relativamente indipendente. Se i Parantropi hanno subito una specializzazione alimentare e morfologica, le specie del primo gruppo si sono evolute mantenendo maggiormente la struttura di base delle forme a loro precedenti. Una delle specie o delle popolazioni che faceva parte di questa variabilità (e non necessariamente una forma di quelle attualmente conosciute nel record fossile) ha fornito il materiale genetico per l'evoluzione del genere umano. Le ultime specie di Australopitechi conosciute (alcuni Parantropi) si sono estinte probabilmente intorno a un milione di anni fa.
Il genere Homo
Le più antiche evidenze di produzione di manufatti litici mediante scheggiatura intenzionale sono le cosiddette industrie a choppers, trovate in Africa orientale (industria olduvaiana). La pietra è lavorata in modo da ottenere superfici taglienti, in genere senza però una particolare rifinitura. Studi più recenti hanno comunque indicato una notevole variabilità di questa industria, che prevede in alcuni casi anche strumenti con una maggiore elaborazione. Per cause di limitata conservazione, è arduo valutare per periodi così antichi la frequenza di utilizzo di strumenti eventualmente prodotti con materiali deperibili (legno, osso). Le forme fossili ipoteticamente associate a questa industria furono conseguentemente chiamate H. habilis. A oggi lo scenario è più ricco, ma meno chiaro. Questi Ominidi vissero intorno ai 2 m.a. fa e avevano una capacità cranica di 600÷800 cm3. In realtà, i fossili inclusi in questo gruppo rappresentano almeno due specie differenti, poco documentate e da alcuni autori neppure incluse nel genere umano. Potrebbero essere infatti forme molto tardive e specializzate di Australopitecine, non necessariamente associate all'evoluzione di specie successive.
Nel periodo immediatamente successivo (da circa 1,9 m.a. dal presente) troviamo però, sempre in Africa orientale, forme che esprimono pienamente la morfologia del genere Homo. Lo scheletro postcraniale indica una postura integralmente bipede, senza traccia di adattamenti locomotori che non siano quelli presenti nell'uomo moderno. Queste forme, oggi identificate con il nome di H. ergaster, si possono ritenere le prime certamente e pienamente umane. Con una capacità cranica media di 800 cm3, queste popolazioni sono associate almeno a partire da 1,6 m.a. fa a un'industria differente da quella a choppers. La pietra veniva lavorata secondo sequenze operative standardizzate e finalizzate all'ottenimento di forme ben precise. Gli strumenti tipici di questa industria (industria acheuleana), denominati bifacciali o amigdale, hanno una qualità che indica un certo livello di astrazione e, se vogliamo, di estetica: la simmetria.
La complessità comportamentale delle prime forme umane le rende più abili nel gestire gli ambienti e le risorse, e in tempi brevissimi le porta ad allargare il proprio areale popolando gli altri continenti. Già 1,7 m.a. fa le troviamo nel Caucaso, e successivamente raggiungono il Sud-Est asiatico (Giava) e l'Estremo Oriente (Pechino). Le forme asiatiche sono più robuste di quelle africane, hanno una capacità cranica di circa 1000 cm3, presentano particolarità che le distinguono morfologicamente e non utilizzano utensili acheuleani. Queste forme, inizialmente chiamate Pitecantropi (uomo di Giava) e Sinantropi (uomo di Pechino), furono poi riunite nella specie H. erectus. All'inizio anche le forme africane furono incluse in questa specie, fino a quando non vennero evidenziate differenze che indicano da un lato una probabile specializzazione delle forme asiatiche (H. erectus), dall'altro un ruolo delle forme africane (H. ergaster) per l'evoluzione delle specie successive. Le differenze tra i due gruppi geografici mostrano comunque spesso una certa gradualità sia nel tempo che nello spazio, tanto da mantenere vivo il dibattito sulla loro effettiva separazione evoluzionistica e di nomenclatura. Tra 2 e 1 m.a. dal presente il popolamento africano rimane comunque poco documentato, con reperti interessanti ma troppo dispersi nel tempo e nello spazio. Intorno a un milione di anni fa queste forme arcaiche di origine africana popolano anche l'Europa, dove fossili come quello rinvenuto in Italia a Ceprano (Frosinone) suggeriscono anche in questo caso una caratterizzazione geografica locale, che viene attualmente indicata (seppure ancora scarsamente conosciuta) col nome di H. antecessor.
Nel periodo successivo (Pleistocene medio, 700.000-120.000 anni) le storie dei tre continenti si separano più o meno gradualmente. In Asia si continuano a trovare resti di H. erectus fino a quasi 200.000 anni, con i recenti ritrovamenti dell'isola di Flores che indicano ipotetiche popolazioni arcaiche residue fino a epoche molto recenti (10.000 anni). Il popolamento afroeuropeo si caratterizza invece per forme molto robuste e più encefalizzate, generalmente ascritte alla specie H. heidelbergensis. In realtà questa specie è più adatta per indicare le popolazioni europee, tra cui quelle dell'eccezionale giacimento di Sima de los Huesos nella Sierra di Atapuerca, in Spagna, che ha restituito i resti di decine di individui. Il record fossile africano è meno ricco e forse più eterogeneo, e viene a volte distinto da quello europeo attraverso la denominazione di H. rhodesiensis. Queste forme afroeuropee hanno una capacità cranica di circa 1200 cm3, usano un'industria di tipo acheuleano e presentano una tipica robustezza delle strutture ossee del cranio, soprattutto nei distretti facciali. La differenza tra forme africane ed europee, al di là del loro riconoscimento come specie differenti o come varietà geografiche di una stessa specie, è importante soprattutto per le rispettive conseguenze evolutive. Tra le forme africane vi erano infatti quelle popolazioni che successivamente avrebbero generato l'uomo anatomicamente moderno (H. sapiens), mentre le popolazioni europee subirono una trasformazione più o meno graduale che sarebbe terminata nella caratterizzazione delle forme neandertaliane (H. neanderthalensis).
Encefalizzazione estrema
I processi di encefalizzazione si riferiscono a un aumento del volume cerebrale rispetto al volume corporeo totale. In paleontologia indici di encefalizzazione sono raramente disponibili con precisione, per mancanza di dati riguardo alla capacità cranica stessa o alla massa corporea. Ma se un certo processo di encefalizzazione è con buona certezza avvenuto nelle forme afroeuropee del Pleistocene medio, in quelle successive (Pleistocene superiore) lo sviluppo volumetrico cerebrale è comunque ancora più evidente.
Sebbene il record fossile sia piuttosto limitato in Africa tra 500.000 e 200.000 anni fa, successivamente a questo periodo troviamo i resti fossili di popolazioni che possiamo definire moderne (H. sapiens). Alcuni reperti come quelli di Gebel Ihroud in Marocco presentano caratteristiche arcaiche che ne rendono incerta l'interpretazione; ma altri resti, come quelli di Omo Kibish in Etiopia, suggeriscono che le forme moderne fossero già distribuite almeno in Africa orientale tra 200.000 e 150.000 anni fa. Intorno a 100.000 anni fa troviamo resti nel Vicino Oriente che, per quanto robusti nell'apparenza, sono assolutamente ascrivibili a quello che viene definito uomo anatomicamente moderno. Dall'altro lato del Mediterraneo, invece, le forme di H. heidelbergensis presentavano variazioni di caratteri che già 120.000 anni fa erano sufficienti per cambiare il loro nome in H. neanderthalensis. Questa specie, distribuita in Europa, Vicino Oriente e Asia occidentale, si presenterà nella sua forma più conosciuta e caratterizzata intorno a 50.000 anni fa.
Le forme moderne e quelle neandertaliane condividono una capacità cranica confrontabile (in media circa 1450 cm3) e un'industria chiamata musteriana, definita da elementi litici molto raffinati ed eterogenei. Le differenze però sono sostanziali, soprattutto se consideriamo le informazioni morfologiche. A parte molte diversità nelle proporzioni o strutture corporee, il cranio presenta due modelli profondamente differenti. Le forme neandertaliane avevano un neurocranio grande ma relativamente arcaico nella struttura, mentre i distretti facciali (denti, strutture nasali, area orbitale, mandibola, ecc.) erano molto specializzati e caratteristici. Le forme moderne presentano al contrario un'assenza di specializzazione, evidente nelle aree facciali, ma una netta ristrutturazione del neurocranio, che coinvolge direttamente la morfologia cerebrale. In queste forme, infatti, l'encefalo non è solo più grande rispetto alle forme del Pleistocene medio, ma presenta anche caratteristiche peculiari che vanno da uno sviluppo eccezionale delle aree parietali a un probabile aumento della vascolarizzazione della corteccia cerebrale.
Quando le forme moderne arrivano in Europa, circa 40.000 anni fa, esse presentano una nuova cultura, la cultura aurignaziana, molto più complessa di quella musteriana e soprattutto associata a componenti astratte e rituali, come pitture rupestri o altre produzioni artistiche. Dopo un periodo di quasi 10.000 anni di convivenza (o comunque di vicinanza) tra queste due specie le forme neandertaliane si estinguono, con le ultime enclaves localizzate nella Spagna meridionale. L'uomo moderno raggiunge l'Oriente intorno ai 50.000 anni, il continente australiano intorno ai 40.000 anni e quello americano probabilmente intorno ai 15.000 anni dal presente. Queste ultime due fasi migratorie suggeriscono un'elevata capacità di organizzazione di queste popolazioni, in grado di gestire spostamenti geografici rilevanti come quelli per mare (nel caso dell'Australia) o attraverso i territori dell'estremo Nord-Est del continente asiatico (nel caso dello Stretto di Bering e delle Americhe).
Teorie a confronto
L'uomo è un primate, quindi un mammifero, quindi un vertebrato, e così via. Le regole della trasmissione genetica, in accordo con le teorie dell'evoluzione darwiniana e della selezione naturale, sono valide anche per il genere umano. La principale difficoltà nell'applicare integralmente le leggi della selezione al genere Homo risiede nelle capacità addizionali di questo gruppo zoologico: la cultura e i suoi particolari adattamenti. La cultura viene definita come adattamento superorganico, e ha la peculiarità di trasmettersi non secondo le regole darwiniane/mendeliane ma piuttosto secondo quelle di Lamarck, ossia per trasmissione di caratteri acquisiti durante la vita di un individuo. L'interazione tra adattamenti darwiniani organici e adattamenti lamarckiani superorganici è alla base dell'ipotesi di una interazione tra queste due componenti, che si potenziano a vicenda: la biologia influenza la cultura, la cultura influenza la biologia. L'encefalizzazione potrebbe di fatto essere il risultato di un processo di questo tipo, ovvero di una 'autocatalisi'. Questo circuito rende a volte le regole dell'evoluzione zoologica non pienamente applicabili all'evoluzione umana. È stato spesso sottolineato, per esempio, come la cultura potrebbe essere un fattore di coesione talmente forte da non permettere ai gruppi umani di diversificarsi fino alla speciazione.
Questo dibattito, insieme ad altre componenti di ordine storico e a volte politico, è stato alla base delle due principali teorie sull'evoluzione del genere Homo, in particolare sull'origine dell'uomo moderno. La teoria della sostituzione (conosciuta anche con la denominazione di Out of Africa) interpreta le varie forme continentali come il risultato di successive sostituzioni da parte di forme che evolvevano in Africa e si spostavano nelle altre regioni già popolate da altre specie. Così H. sapiens si origina in Africa, si sposta in Europa sostituendo i Neandertaliani e in Asia sostituendo H. erectus. È molto importante notare che una sostituzione del genere non comporta necessariamente una 'superiorità' adattativa delle forme moderne, ma può anche essere il risultato di un'estinzione autonoma delle specie locali e di un successivo spostamento in territori dunque liberi delle nuove specie emigranti. Al contrario, la teoria multiregionale ritiene che le differenze tra forme arcaiche e forme moderne non siano tali da giustificare una separazione filogenetica e suggerisce che l'evoluzione umana sia stata il risultato di un cambiamento globale senza distinzioni geografiche nette, o comunque riconoscibili in termini paleontologici.
Il dibattito tra queste due teorie è sempre vivo e spesso condotto su un piano non propriamente scientifico. Spesso, per esempio, le due teorie vengono prese in esame per confutare il concetto di razza nella specie umana, quando invece entrambe sono compatibili sia con il concetto di razza sia con la sua negazione. Non mancano inoltre tentativi di combinazione delle due teorie, secondo i quali nuove forme avrebbero colonizzato nuovi territori assorbendo geneticamente le popolazioni residenti. Il punto fondamentale è che il record fossile è in genere troppo frammentato nel tempo, nello spazio e nella morfologia per permettere un approccio statistico e formale che possa falsificare una teoria a favore di un'altra. In Europa le evidenze di una sostituzione sono piuttosto solide. Non vi sono attestazioni di continuità o mescolamento nella morfologia delle forme moderne e di quelle neandertaliane, e anche i processi di sviluppo sembrano distinti. Più recentemente anche i dati molecolari hanno riscontrato una sostanziale separazione della variabilità genetica tra questi due gruppi. Al contrario, in Asia le indicazioni sono più incomplete, le dinamiche del Pleistocene medio in Africa non ancora note e i dati sul continente australiano a volte contraddittori.
In questo tipo di analisi bisogna comunque tener presente che le ipotesi evoluzionistiche rappresentano modelli utili all'interpretazione di una storia che, per quanto ignota e incerta, è comunque avvenuta, ed è soprattutto unica. Con l'incremento del record fossile, l'introduzione di metodologie complementari e l'approccio di paradigmi sperimentali, la paleoantropologia è oggi una disciplina scientifica a tutti gli effetti, sempre più lontana da quella componente narrativa e descrittiva che ne ha decretato da un lato la diffusione mediatica, dall'altro la frequente difficoltà di difendere le sue scoperte e trasformare i suoi contributi in patrimonio comune. Qualsiasi modello potrà utilizzare solo le informazioni e i principî parziali disponibili in un dato momento, e non può essere ritenuto né definitivo né completo. Lo scopo di questi modelli deve essere quindi quello di essere utili all'interpretazione della nostra storia passata, della nostra condizione presente e delle nostre potenzialità future.
Bibliografia
Antón 2003: Antón, Susan C., Natural history of Homo erectus, "Yearbook of physical anthropology", 46, 2003, pp. 126-170.
Bruner 2003: Bruner, Emiliano - Manzi, Giorgio - Arsuaga, Juan Luis, Encephalization and allometric trajectories in the genus Homo: evidence from the Neandertal and modern lineages, "Proceedings of the National Academy of Sciences USA", 100, 2003, pp. 15335-15340.
Finlayson 2005: Finlayson, Clive, Biogeography and evolution of the genus Homo, "Trends in ecology and evolution", 20, 2005, pp. 457-463.
Kay 1997: Kay, Richard F. - Ross, Callum - Williams, Blythe A., Anthropoid origins, "Science", 275, 1997, pp. 797-804.
Krings 2000: Krings, Matthias e altri, A view of Neandertal genetic diversity, "Nature genetics", 26, 2000, pp. 144-146.
McBrearty, Brooks 2000: McBrearty, Sally - Brooks, Alison S., The revolution that wasn't: a new interpretation of the origin of modern human behavior, "Journal of the human evolution", 39, 2000, pp. 453-563.
Rightmire 1998: Rightmire, G. Philip, Human evolution in the Middle Pleistocene: the role of Homo heidelbergensis, "Evolutionary anthropology", 6, 1998, pp. 218-227.
Stringer 2002: Stringer, Chris, Modern human origins: progress and prospects, "Philosophical transactions of the Royal Society of London, B", 357, 2002, pp. 563-579.
Wood, Collard 1999: Wood, Bernard - Collard, Mark, The human genus, "Science", 284, 1999, pp. 65-71.
© Istituto della Enciclopedia Italiana - Riproduzione riservata


