Percezione
Enciclopedia Italiana - VI Appendice (2000)
Percezione
(XXVI, p. 715)
La p. costituisce il principale processo psicologico che presenti un interesse multidisciplinare, dalla filosofia alla psicologia, dalla neurofisiologia all'intelligenza artificiale e alla robotica. Questa posizione privilegiata è direttamente connessa al ruolo che la tradizione filosofica occidentale ha assegnato alla p. in quanto processo fondamentale che consente alla mente di conoscere il mondo esterno. Senza le informazioni relative alla realtà esterna, la mente non potrebbe formare rappresentazioni interne, elaborare concetti e schemi cognitivi e progettare programmi di azione volti a trasformare la realtà stessa. Ogni sistema filosofico che abbia trattato i processi di conoscenza è partito quindi da un esame delle funzioni e delle caratteristiche della p.; molte correnti filosofiche del primo Novecento hanno fatto riferimento alla centralità della p. nello studio dell'uomo (Merleau-Ponty 1945), mentre nelle ricerche più recenti di filosofia della mente la p. rappresenta il tema di avvio della discussione sulle caratteristiche della mente umana (Smith, Jones 1986).
Al di là della convergenza interdisciplinare sul problema della p., che determina la compresenza di procedure e metodi diversi d'indagine, la maggior parte dei ricercatori contemporanei è concorde nel riconoscere la necessità di una verifica empirica, oggettiva, dei fenomeni percettivi studiati. Negli ultimi vent'anni si è peraltro ripresentato il problema del riferimento ai dati soggettivi per studiare la percezione. Infatti, la possibilità di esperire direttamente un dato fenomeno (per es., la costanza del colore degli oggetti nonostante la variazione di luminanza ambientale) ha permesso al ricercatore di essere lui stesso l'osservatore del fenomeno senza ricorrere ad altri osservatori. Anche nel passaggio dalla 'psicologia filosofica' alla 'psicologia scientifica', segnato proprio dalle ricerche sulla sensazione e la p. di metà Ottocento, i ricercatori (come G.T. Fechner) erano allo stesso tempo gli osservatori dei fenomeni indagati. Quindi, il percetto è stato studiato come fenomeno nel senso etimologico della parola ('ciò che appare'); molte ricerche di psicologia della p. sono condotte ancora oggi con il metodo fenomenologico: lo psicologo studia il fenomeno percettivo quale lo esperisce egli stesso.
È chiaro che ciò favorisce la diffusione di una varietà e molteplicità di risultati, perché ciascun osservatore esperisce direttamente i fenomeni illustrati (come nel caso di molte illusioni ottiche raffigurate comunemente nei manuali di psicologia). Il metodo fenomenologico segue, comunque, procedure rigorose di manipolazione delle variabili (per cui si parla, più propriamente, di metodo fenomenologico sperimentale). Questa metodologia è stata adottata in particolare dalla teoria della forma (Gestalttheorie) che ha orientato le ricerche sulla p. dagli anni Dieci fino agli anni Cinquanta circa del Novecento (Kanizsa, Caramelli 1988). Negli anni Sessanta gli studi sulla p. hanno assunto un'impostazione teorica e metodologica diversa, centrata sull'adozione dell'orientamento cognitivista. Il cognitivismo sostituì alla concezione della mente come 'scatola nera' (black box), struttura indifferenziata di stimoli in entrata e di risposte in uscita, una teoria più articolata e complessa per la quale la mente è composta da sottostrutture specializzate in operazioni cognitive distinte (p., attenzione, memoria ecc.), tra loro comunicanti e interagenti. Il libro di D.E. Broadbent, Perception and communication (1958), come indica lo stesso titolo, legava strettamente il processo della p. a quello della comunicazione e trasmissione dell'informazione all'interno della mente, in una stretta relazione con altri processi cognitivi, in primo luogo l'attenzione e la memoria. Da allora, la p. è considerata un processo guidato da altri processi cognitivi o, in altri termini, come un'operazione flessibile di 'cattura' ed esplorazione attiva dell'informazione esistente nel mondo esterno, in funzione delle aspettative e delle esperienze pregresse dell'organismo. Al contrario, in altre teorie ricollegantisi alla teoria della forma, la p. è concepita come un processo autonomo che ha leggi proprie ed è relativamente indipendente dagli altri processi mentali (Kanizsa 1980, 1991).

Mentre la teoria della forma riteneva che le leggi dell'organizzazione percettiva (le leggi gestaltiche) avessero un fondamento neurofisiologico, le teorie contemporanee di orientamento cognitivista hanno distinto tra un livello strettamente fisiologico e un livello psicologico della percezione. Il primo livello può essere considerato quello trattato tradizionalmente come 'sensazione', relativo all'architettura funzionale dei neuroni e alle leggi della psicofisica regolanti l'elaborazione degli attributi fisici degli stimoli (intensità, lunghezza d'onda ecc.); nel secondo livello, invece, rientra la problematica relativa all'elaborazione cognitiva dell'informazione. Questa distinzione può essere illustrata con l'esempio della fig. 1. A livello fisiologico, si tratta di un insieme di punti, linee e macchie disposte su uno spazio bidimensionale, che vengono elaborate dal sistema visivo rispetto a parametri fisici relativi (intensità, contrasto, grandezza ecc.); a livello di p., si tratta invece di un 'percetto', un'interpretazione cognitiva di ciò che è stato elaborato dal sistema sensoriale della visione. L'informazione cognitiva trasmessa dall'espressione cane dalmata consente di interpretare tale informazione puramente fisica. Questa relazione tra il livello fisico e il livello cognitivo è stata posta come interazione tra quelli che sono stati definiti processi bottom-up e top-down (Lindsay, Norman 1972). L'elaborazione bottom-up è basata su informazioni e indizi presenti nella realtà, che pervengono direttamente ai sistemi sensoriali (le macchie dell'esempio precedente). Sebbene tale informazione, per essere interpretata e riconosciuta come un percetto significativo, necessiti dell'interazione con il processo top-down (l'informazione verbale), è evidente che gli indizi sensoriali non possono scendere al di sotto di una soglia che consenta tale interazione. Studi recenti hanno messo in evidenza che questo interscambio tra i due tipi di elaborazione avviene al livello inconscio e che il riconoscimento di una figura frammentata, come nell'esempio di cui sopra, è segnalato dall'attività elettrica cerebrale prima che l'osservatore affermi di aver riconosciuto la figura (Viggiano, Kutas 1998). Un'altra problematica degli studi attuali sulla p. è, quindi, la compresenza di componenti consce e inconsce (argomento che rientra nel tema più ampio dell'inconscio cognitivo: Conte, Gennaro 1989).
Sebbene l'interpretazione del percetto richieda il confronto con l'informazione passata e, quindi, il processo della p. sia strettamente legato ai processi della memoria, rimane aperta la questione del fenomeno della costruzione di un percetto al livello fisiologico. In altri termini, anche se non si ha a disposizione l'informazione top-down per riconoscere nella fig. 1 un cane dalmata, ciò che si vede è comunque un percetto con la sua organizzazione spaziale, le sue caratteristiche cromatiche ecc. Per la comprensione di questo livello di analisi del processo percettivo, un contributo importante è stato dato dalla neurofisiologia a partire dalla fine degli anni Cinquanta. Le ricerche di V.B. Mountcastle sulla corteccia somatosensoriale, di D.H. Hubel e T.N. Wiesel sulla corteccia visiva e di Y. Katsuki e collaboratori sulla corteccia uditiva dimostrarono l'esistenza di un'architettura modulare della corteccia sensoriale: nell'ambito di ciascuna modalità sensoriale (somatosensoriale, visiva o uditiva) vi sono colonne o moduli distinti di neuroni, e ciascuna colonna è selettiva per determinati parametri dell'informazione in arrivo. Per due attributi fondamentali dell'informazione visiva, la distribuzione della luminanza nello spazio (o frequenza spaziale) e l'orientamento dello stimolo, sono stati individuati nella corteccia visiva (nelle aree 17, 18 e 19; attuale nuova denominazione: aree V1-V5) neuroni altamente specializzati, sensibili a particolari frequenze spaziali od orientamenti (per queste ricerche, condotte negli anni Sessanta e Settanta: Maffei, Mecacci 1979; Hubel 1988).
La p. di uno stimolo visivo, per es. un quadrato, avrebbe avuto come fondamento l'integrazione tra le risposte di vari neuroni selettivi a differenze di luminanza orientate nello spazio (i lati neri, orizzontali e verticali, del quadrato sullo sfondo bianco), secondo un modello dell'interazione neuronale già proposto da D.O. Hebb (1949) e ripreso da Hubel e Wiesel. Negli anni Ottanta fu proposta una nuova classificazione alla quale hanno fatto riferimento le ricerche contemporanee sulle basi neuronali della p. visiva. M.S. Livingstone e D.H. Hubel (1984) hanno distinto due sistemi visivi principali: il sistema magnocellulare e il sistema parvocellulare. Il sistema magnocellulare ha origine nelle cellule gangliari retiniche Y o M, sensibili alle differenze di luminanza che colpiscono il proprio campo recettivo (la porzione di campo visivo esterno cui risponde la cellula) e non alle differenze di lunghezza d'onda (colore); si connette sia agli strati magnocellulari del corpo genicolato laterale (così chiamati per la grande dimensione delle cellule), sia ai collicoli superiori, e termina in aree corticali distinte lungo la via dorsale (fig. 2). Il sistema parvocellulare ha origine nelle cellule gangliari retiniche X o P, sensibili al colore e non sensibili alle differenze di luminanza; si connette agli strati parvocellulari del corpo genicolato laterale (così chiamati per la piccola dimensione delle cellule) e termina in altre aree corticali lungo la via ventrale (fig. 2). Alcune regioni corticali di alta densità (intensa colorazione messa in evidenza con la tecnica della citocromo-ossidasi), dette blobs, comprendono cellule altamente selettive al colore, mentre le cellule localizzate nelle regioni con bassa densità, dette interblobs, rispondono selettivamente all'orientamento e alla forma degli stimoli. I due sistemi avrebbero funzioni diverse nella p. visiva: il sistema parvocellulare sarebbe impegnato nella p. del contrasto (differenza di luminanza nello spazio) e del movimento (la via dorsale ha connessioni con la regione dei lobi temporali, nella quale risiedono i neuroni che presiedono al movimento), mentre il sistema magnocellulare elaborerebbe in modo specializzato la forma e il colore. Secondo la classificazione di L.G. Ungerleider e M. Mishkin (1982), la via dorsale sarebbe dedicata all'elaborazione degli attributi degli stimoli visivi (grandezza, orientamento, forma e colore), cioè, in sintesi, alla dimensione 'che cosa è?' nella p. di uno stimolo. La via dorsale sarebbe invece dedicata alla localizzazione spaziale dello stimolo o alla dimensione 'dov'è?' Secondo M.A. Goodale e A.D. Milner (1992) la differenziazione funzionale tra le due vie visive è ancora più generale: la via dorsale riguarderebbe l'analisi degli stimoli visivi in funzione dei movimenti che l'osservatore deve compiere per interagire con essi, mentre la via ventrale sarebbe implicata nella rappresentazione cognitiva degli stimoli stessi (la dicotomia è quindi tra azione e cognizione nel processo percettivo).
La componente top-down della p. viene ricondotta alla capacità della mente di formarsi rappresentazioni interne degli stimoli esterni. Quando un osservatore è informato che lo stimolo della fig. 1 è un cane dalmata, egli può utilizzare questa informazione per riconoscere tale stimolo solo se nella sua memoria è già presente la rappresentazione del cane dalmata. Rappresentazioni di questo tipo sono dette anche prototipi per indicare che esse sono una descrizione generale di uno stimolo o di una determinata classe di stimoli (cane dalmata, cane, animale) rispetto a esemplari specifici (il cane dalmata abbozzato nella fig. 1, quello che l'osservatore può aver visto ai giardini pubblici, il proprio cane dalmata ecc.).
Le teorie sulla formazione dei prototipi sono molteplici, ma possono essere raccolte in quattro grandi gruppi: teorie della sagoma, teorie dell'analisi delle caratteristiche, teorie strutturali e teorie computazionali (Bruce, Green, Georgeson 1985, 1996²).
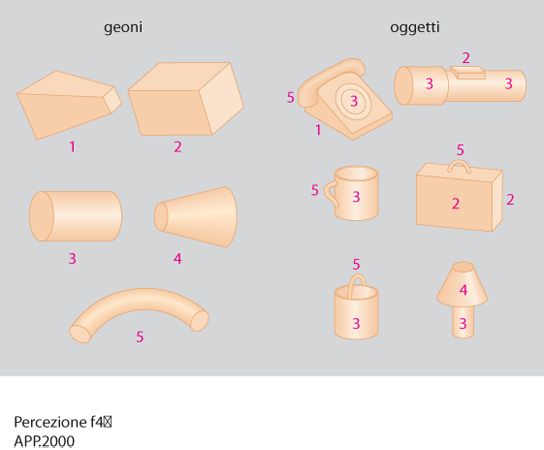
Per le teorie della sagoma uno stimolo è riconosciuto quando è realizzato con successo il confronto (template matching) con una versione standard dello stimolo depositata nella memoria dell'osservatore. Secondo le teorie delle caratteristiche, la mente opera un'analisi (feature analysis) di alcuni elementi essenziali dello stimolo (per es., per la lettera P sono un segmento verticale e un semicerchio), e confronta il risultato dell'analisi con una sorta di inventario depositato in memoria (per cui tutti gli stimoli che presentano un segmento verticale e un semicerchio sono identificati come una P). Queste prime due teorie furono molto diffuse negli anni Sessanta e Settanta (Selfridge 1959; Neisser 1967; Lindsay, Norman 1972). In seguito, sono state avanzate altre teorie più complesse come le teorie strutturali (tra cui quella di Winston 1977, 1984²) che si fondano più sulle relazioni tra gli elementi che sul catalogo degli elementi presenti nello stimolo (per es., è la relazione tra i lati di un triangolo che permette di identificarlo come tale più che la presenza di tre segmenti, due obliqui e uno orizzontale). Le teorie strutturali hanno permesso di descrivere sotto forma di algoritmi, da implementare in un programma di simulazione al calcolatore, le regole di generazione e riconoscimento di uno stimolo visivo e sono confluite nelle teorie computazionali, tra le quali la più innovativa è stata quella di D. Marr (1982). Le teorie computazionali indicano quali sono le componenti essenziali o primitive e le regole che portano al riconoscimento di uno stimolo (per es., nella descrizione fondata sul modello tridimensionale o 3-D di Marr, Nishihara 1978; fig. 3). Anche la teoria di I. Biederman (1987, 1990) descrive componenti primitive (solidi detti geoni; fig. 4) sulla cui base, e attraverso regole determinate, possono essere composti i singoli oggetti. Nell'approccio connessionistico le teorie strutturali e computazionali sono state sottoposte a verifica addestrando le reti neurali artificiali a riconoscere stimoli relativamente complessi (McClelland, Rumelhart 1985; Churchland, Sejnowski 1992; Hummel, Biederman 1992).
La p. uditiva è stata studiata soprattutto al livello dello stadio sensoriale di analisi degli attributi (intensità, frequenza, ritmo ecc.) di uno stimolo uditivo. Le ricerche di psicofisica nell'uomo e di elettrofisiologia negli animali hanno dimostrato la selettività dei neuroni della corteccia uditiva per tali parametri (Kandel, Schwartz 1981, 1985²). Lo stadio di elaborazione cognitiva dell'informazione uditiva è stato indagato soprattutto per quanto riguarda il riconoscimento di suoni linguistici e dell'informazione verbale (fonemi, parole) sia in soggetti normali, sia in pazienti cerebrolesi (Denes, Semenza, Magno Caldognetto 1990, 1996²). I modelli adottati sono simili a quelli descritti per la p. visiva, con la distinzione in stadi di elaborazione e il confronto tra lo stimolo in entrata e la traccia prototipica depositata in memoria. Un altro settore di ricerca riguarda la p. della musica, in particolare nelle sue componenti strutturali di melodia e ritmo. Dopo le ricerche classiche della psicofisica e della teoria della forma, anche in questo settore sono stati adottati modelli cognitivistici (Slaboda 1985).
Negli ultimi vent'anni del Novecento si è rafforzata l'interazione tra psicologia e altre aree, come quelle dell'intelligenza artificiale, dell'informatica e dell'ingegneria cognitiva, allo scopo di produrre macchine capaci di percepire e identificare stimoli visivi e uditivi. Sebbene la realizzazione di queste macchine artificiali abbia un esplicito fine applicativo, essa consente anche una più approfondita conoscenza dei meccanismi e dei processi della p. naturale. I modelli connessionistici, fondati sull'analisi in parallelo degli stimoli, permettono di sviluppare programmi su calcolatore più flessibili e sofisticati. Il problema se una macchina artificiale che percepisce sia simile a una macchina 'naturale' (Poggio 1984) è stato ampiamente dibattuto all'interno degli studi sull'intelligenza artificiale e delle scienze cognitive (Cordeschi 1996) ed è stato spesso posto nei termini funzionalistici di H. Putnam (1975): occorre determinare quali siano i processi che consentono la p. indipendentemente dal supporto materiale (elettronico, nel calcolatore, o biologico, nel cervello) nel quale avviene l'elaborazione.
Alcuni psicologi (Bruce, Green, Georgeson 1985, 1996²) hanno impostato la discussione sulle caratteristiche generali dei processi percettivi di un osservatore naturale contrapponendo teorie indirette, del tipo delle precedenti, e teorie dirette. Il principale esponente delle teorie dirette è stato J.J. Gibson (1966, 1979), che ha rifiutato le concezioni tradizionali della p. e ha proposto un approccio ecologico. Secondo Gibson, un osservatore 'cattura' direttamente l'informazione visiva qual è data nel proprio ambiente e in funzione delle azioni che deve compiere. Non vi sono, quindi, stadi successivi di analisi che conducono al percetto finale, a partire da elementi separati, elaborati uno per uno e poi ricomposti in unità superiori. Questa impostazione elementistica produce una frantumazione della p. in stadi e operazioni distinte (è l'effetto delle condizioni artificiali di laboratorio), ma non permette di conoscere i processi reali della percezione. Con riferimento alla teoria diretta di Gibson sono stati compiuti vari esperimenti di confronto tra condizioni percettive realizzate in laboratorio e in ambiente naturale (per es., si osserva un oggetto e si deve prevedere il tempo di collisione con esso, come durante la guida di un'automobile: Tresilian 1995), ottenendo risultati differenti a seconda delle condizioni, artificiali o ecologiche. In questa prospettiva, il processo percettivo è considerato nel ruolo che svolge all'interno dei meccanismi di controllo del movimento e dell'azione (Turvey 1977; Bruce, Green, Georgeson 1985, 1996²) più che come sistema di osservazione passiva della realtà esterna.
bibliografia
M. Merleau-Ponty, Phénoménologie de la perception, Paris 1945 (trad. it. Milano 1965).
D.O. Hebb, The organization of behavior, New York 1949 (trad. it. Milano 1975).
D.E. Broadbent, Perception and communication, New York 1958.
O.G. Selfridge, Pandemonium. A paradigm for learning, in The mechanisms of thought processes, London 1959.
J.J. Gibson, The senses considered as perceptual systems, Boston 1966.
U. Neisser, Cognitive psychology, New York 1967 (trad. it. Firenze 1976).
N. Pastore, Selective history of theories of visual perception 1650-1950, New York 1971.
P.H. Lindsay, D.A. Norman, Human information processing. An introduction to psychology, New York 1972 (trad. it. L'uomo elaboratore di informazioni. Introduzione cognitiva alla psicologia, Firenze 1983).
H. Putnam, Mind, language and reality, Cambridge-New York 1975 (trad. it. Milano 1987).
M.T. Turvey, Contrasting orientations to the processing of visual information, in Psychological review, 1977, pp. 67-89.
P.H. Winston, Artificial intelligence, Reading (Mass.) 1977, 1984².
D. Marr, H.K. Nishihara, Representation and recognition of the spatial organization of a three-dimensional shape, in Proceedings of the Royal society of London, 1978, 200, pp. 269-94.
J.J. Gibson, The ecological approach to visual perception, Boston 1979.
L. Maffei, L. Mecacci, La visione. Dalla neurofisiologia alla psicologia, Milano 1979.
G. Kanizsa, Grammatica del vedere. Saggi su percezione e Gestalt, Bologna 1980.
E.R. Kandel, J.H. Schwartz, Principles of neural science, New York 1981, 1985² (trad. it. Milano 1988).
D. Marr, Vision, a computational investigation into the human representation and processing of visual information, San Francisco 1982.
L.G. Ungerleider, M. Mishkin, Two cortical visual systems, in Analysis of visual behaviour, ed. D.J. Ingle, M.A. Goodale, R.J.W. Mansfield, Cambridge (Mass.) 1982, pp. 549-86.
M.S. Livingstone, D.H. Hubel, Anatomy and physiology of a colour system in the primate visual cortex, in Journal of neuroscience, 1984, 4, pp. 309-56.
T. Poggio, La visione nell'uomo e nella macchina, in Le Scienze, 1984, 190.
V. Bruce, P.R. Green, M.A. Georgeson, Visual perception. Physiology, psychology and ecology, Hove 1985, 1996².
J.L. McClelland, D.E. Rumelhart, Distributed memory and the representation of general and specific information, in Journal of experimental psychology. General, 1985, 114, pp. 159-88.
J.A. Slaboda, The musical mind, Oxford-New York 1985 (trad. it. Bologna 1988).
P. Smith, O.R. Jones, The philosophy of mind, Cambridge 1986.
I. Biederman, Recognition by components. A theory of human image understanding, in Psychological review, 1987, 94, pp. 115-47.
D.H. Hubel, Eye, brain and vision, New York 1988 (trad. it. Bologna 1989).
G. Kanizsa, N. Caramelli, L'eredità della psicologia della Gestalt, Bologna 1988.
M. Conte, A. Gennaro, Inconscio e processi cognitivi, Bologna 1989.
I. Biederman, Higher-level vision, in An invitation to cognitive science, ed. D.N. Osherson, S.M. Kosslyn, J.M. Hollerbach, 2° vol., Visual cognition and action, Cambridge (Mass.) 1990, pp. 41-72.
F. Denes, C. Semenza, E. Magno Caldognetto, Disturbi fonologici nell'afasia, in Manuale di neuropsicologia, a cura di G. Denes, L. Pizzamiglio, Bologna 1990, 1996², pp. 258-87.
G. Kanizsa, Vedere e pensare, Bologna 1991.
P.S. Churchland, T.J. Sejnowski, The computational brain, Cambridge (Mass.) 1992 (trad. it. Bologna 1996).
M.A. Goodale, A.D. Milner, Separate visual pathways for perception and action, in Trends in neuroscience, 1992, 15, pp. 20-25.
J.E. Hummel, I. Biederman, Dynamic binding in a neural network for shape recognition, in Psychological review, 1992, pp. 480-517.
J.R. Tresilian, Perceptual and cognitive processes in time-to-contact estimation: analysis of prediction-motion and relative judgment tasks, in Perception and psychophysics, 1995, 57, pp. 231-45.
R. Cordeschi, L'intelligenza artificiale, in L. Geymonat, Storia del pensiero filosofico e scientifico, 8° vol., tomo 3°: Il Novecento, a cura di E. Bellone, C. Mangione, Milano 1996, pp. 145-200.
M.P. Viggiano, M. Kutas, The covert interplay between perception and memory: an event-related potential evidence, in Electroencephalography and clinical neurophysiology, 1998, 108, pp. 435-39.


