Selezione
Enciclopedia on line
Operazione di scelta finalizzata a trarre da un gruppo gli elementi migliori o più adatti a determinati scopi.
Agraria
S. delle sementi Processo mediante il quale le sementi sono liberate dalle impurità, dai semi estranei e dai semi rotti. Essiccate, quindi immesse in contenitori o in sili allo stato sfuso, le sementi passano dai contenitori o dai sili alla tramoggia di carico dell’impianto di s., nel quale, preventivamente, attraversano una tarara che elimina mediante ventilazione le impurità leggere; successivamente, passano in un complesso costituito da un concentratore per la classificazione in funzione della densità, da un separatore a stacci per l’eliminazione di impurità grosse e fini e da due svecciatoi per l’eliminazione di corpi allungati (semi, avena, orzo ecc.) e di corpi tondeggianti (veccia, grani grossi ecc.).
Biologia
S. naturale La principale forza che modifica le frequenze alleliche all’interno di una popolazione in risposta a precise esigenze di adattamento all’ambiente circostante, rappresentando quindi uno dei più importanti processi che causano cambiamenti evolutivi.
Cenni storici
I primi a identificare la s. come causa principale dell’evoluzione biologica e a indicarla come forza guida dell’adattamento e della variabilità delle popolazioni furono due naturalisti inglesi, A.R. Wallace e C. Darwin, con una comunicazione congiunta presentata alla Linnean Society di Londra il 1° luglio 1858. Per l’elaborazione di questa rivoluzionaria teoria, per entrambi i ricercatori fu determinante la lettura critica dell’opera di T.R. Malthus del 1798 An essay on the principle of population e del trattato di C. Lyell del 1830 Principles of geology. Dall’osservazione che in natura il numero delle popolazioni rimane relativamente costante, malgrado tutti gli organismi siano in grado di riprodursi in maniera geometrica, si dedusse che doveva esistere una qualche forma di competizione per la vita. Wallace intuì che la sostanziale costanza numerica delle popolazioni naturali fosse da ricondurre all’azione stabilizzante della s. naturale, che consente solo a pochi di sopravvivere e riprodursi in presenza di pressioni ambientali, tra cui, in maniera determinante a suo giudizio, la competizione per le risorse. Darwin, come si apprende dalla sua autobiografia, fu particolarmente colpito dalla citazione di Malthus secondo cui nella «lotta per la vita... le variazioni vantaggiose tendono a essere conservate e quelle sfavorevoli a essere distrutte». La grande novità della concezione darwiniana riguardo alla s. sta nell’idea che essa non si limita a essere un processo eliminativo, ma può anche consentire l’insorgere di nuovi caratteri utili per la popolazione. Nella sua opera fondamentale, On the origin of species pubblicata nel 1859, Darwin, partendo dall’osservazione che esiste una naturale variabilità morfologica tra gli individui all’interno della stessa specie, formulò la sua teoria sulla s. naturale in base alla quale «ogni essere vivente, per quanto trascurabile sia la sua diversità, se essa in qualche modo rappresenta un vantaggio per esso, ... allora avrà una migliore opportunità di sopravvivere» e precisò il suo concetto puntualizzando che è importante non solo la sopravvivenza dell’individuo, ma il successo biologico della sua progenie.
L’intuito di Darwin acquista un particolare rilievo considerando che quando delineò la sua teoria non erano ancora note le basi molecolari della variabilità fra gli individui di una popolazione, ossia le mutazioni. Solo successivamente, con la dimostrazione della base genetica delle variazioni ereditarie e con il delinearsi del concetto di geni, mutazioni e alleli, la logica innovativa della sua teoria fu confermata.
Definizione e descrizione
Nello studio dell’evoluzione bisogna tenere presente che l’oggetto di questo processo non è il singolo individuo ma la popolazione nel suo insieme; si devono cioè prendere in considerazione le frequenze alleliche e genotipiche delle popolazioni piuttosto che i meccanismi ereditari all’interno di una famiglia, ed è pertanto necessario inquadrare lo studio della s. e quindi dell’evoluzione nel contesto della genetica di popolazione. Le mutazioni e le migrazioni introducono nuovi alleli nelle popolazioni, ma è la s. che modifica le frequenze alleliche all’interno di esse in risposta a specifiche esigenze ambientali. In ogni popolazione, a un dato momento, sono presenti individui con diversi genotipi, alcuni dei quali più adatti rispetto ad altri alle condizioni ambientali e quindi alla riproduzione. La s. opera infatti in base alla ‘capacità riproduttiva differenziale’ dei diversi genotipi, provocando, nel corso del tempo, cambiamenti anche consistenti delle frequenze alleliche, e questo costituisce un importante meccanismo evolutivo. In base alla legge di Hardy e Weinberg infatti la popolazione è in equilibrio, e quindi non si modifica, quando le frequenze alleliche rimangono costanti, in quanto tutti i genotipi hanno uguali capacità riproduttive e di sopravvivenza (➔ equilibrio). La s. agisce quando gli organismi che hanno una particolare combinazione di genotipo/fenotipo sono competitivi per determinate caratteristiche quali, per es., la fertilità, la longevità, la resistenza alle malattie, le necessità alimentari, l’abilità a difendersi o a nascondersi ai nemici ecc. La probabilità di un particolare fenotipo di sopravvivere e dare progenie è misurata dalla sua fitness (➔), che rappresenta il potenziale riproduttivo totale o efficienza biologica.
Teoria matematica della selezione
Si prenda, come esempio, una popolazione in cui siano presenti i seguenti genotipi AA, Aa, aa. Dal punto di vista matematico la differenza tra fitness di un genotipo in esame rispetto a un altro preso come controllo ottimale è chiamata coefficiente di s. (s). Se per il fenotipo conferito dal genotipo aa, solo 99 individui su 100 sono in grado di riprodursi con successo, allora s=0,01. Nel caso in cui il genotipo aa sia un omozigote letale in un ambiente in cui i due genotipi AA e Aa hanno uguali capacità riproduttive, allora il suo coefficiente di s. è s=1,0 e l’allele a è trasmesso solo dagli eterozigoti (individui portatori). In una popolazione con frequenze iniziali p0 (per l’allele A) e q0 (per l’allele a), con un coefficiente di s. s=1 per il genotipo aa, l’effetto della s. per tutte le successive generazioni sarà calcolato dalla formula:
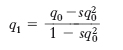
dove n=numero delle generazioni trascorse a partire dai valori iniziali di p0 e q0. Se si prende in considerazione una popolazione con frequenze alleliche p0 (A)=q0(a)=0,5, in cui il genotipo aa ha s=1, alla prima generazione la frequenza dell’allele a verrà ridotta rapidamente, in conseguenza dell’alta percentuale di genotipi aa, e apparirà dimezzata (0,25) solo dopo il passaggio di due generazioni. Dopo sei generazioni la frequenza di a è ulteriormente ridotta della metà (0, 12) e da questo momento in poi la maggior parte degli alleli a nella popolazione è portata dagli eterozigoti. Poiché la s. opera sui fenotipi e non sui singoli alleli del genotipo, gli eterozigoti non sono selezionati contro e pertanto le riduzioni nella frequenza dell’allele a avvengono molto lentamente nel corso delle generazioni successive e solo in relazione alla morte casuale degli eterozigoti. Per questa ragione, fino a quando sono statisticamente possibili unioni tra eterozigoti, è difficile mediante la s. eliminare un allele recessivo dalla popolazione. Quando il coefficiente di s. (s) è minore di 1,0, è possibile calcolare l’effetto della s. in ciascuna delle generazioni successive con qualsiasi valore di s, p0 e q0, usando la formula:
Meccanismi della s. naturale
Il processo di s. può agire solo in presenza di variabilità genetica, cioè se nella popolazione, in conseguenza di un evento di mutazione, sono presenti almeno due alleli diversi per un determinato gene. Una mutazione, a priori, non può essere considerata né vantaggiosa né deleteria; la scelta dipende dall’ambiente in cui si esprime e quindi la stessa mutazione può rappresentare un vantaggio adattativo in un ambiente e uno svantaggio grave in un altro. Una convincente conferma di ciò si ha osservando l’involuzione di molti organi nelle specie parassite, in cui è sicuramente vantaggioso eliminare strutture e quindi funzioni diventate inutili nell’adattamento dell’organismo al nuovo ambiente.
Sono stati studiati molti aspetti della s. sia nelle popolazioni naturali sia in quelle allevate in laboratorio. Un classico esempio di meccanismo di azione della s. in popolazioni naturali è quello descritto per la falena Biston betularia, molto comune in Inghilterra. Prima del 1850, il 99% della popolazione era costituito da falene con ali di colore grigio maculato di bianco, che consentiva all’individuo di mimetizzarsi perfettamente con il colore della corteccia degli alberi coperti dai licheni. Gas tossici prodotti dalle industrie uccisero i licheni e i muschi che crescevano sugli alberi e sulle costruzioni e la fuliggine scurì il paesaggio; le falene con la colorazione chiara divennero un facile bersaglio per i loro predatori, mentre le rare farfalle della stessa specie con il fenotipo mutante ad ali scure immediatamente ebbero un grande vantaggio selettivo in virtù della loro naturale possibilità di mimetizzarsi e quindi di sottrarsi alla predazione. In pochi anni, quindi, si verificò un rapido aumento della frequenza di questo fenotipo, in conseguenza del rapido cambiamento ambientale. Esperimenti di laboratorio hanno dimostrato che il fenotipo ad ali scure è dovuto alla presenza nel genotipo di almeno un singolo allele dominante C, mentre il fenotipo ad ali chiare è l’espressione del genotipo cc. Nel 1964, in seguito all’adozione di leggi severe per l’inquinamento ambientale, intorno all’area di Manchester la frequenza della varietà di falene ad ali chiare ha cominciato ad aumentare, dimostrando la stretta relazione tra ambiente e selezione.
Un esempio analogo nell’uomo è costituito da due malattie ereditarie che interessano le emoglobine e che in alcune popolazioni presentano una elevata incidenza: la talassemia e l’anemia falciforme. Queste malattie rappresentano su scala mondiale le malattie monogeniche più frequenti, seconde forse soltanto alla fibrosi cistica. Gli alleli responsabili di queste emoglobinopatie hanno raggiunto frequenze così alte in quanto conferiscono agli individui eterozigoti (portatori di un solo allele mutato) un’aumentata resistenza a contrarre la malaria a opera di Plasmodium falciparum iniettato nell’uomo dalla puntura della zanzara Anopheles. Questa interazione tra ambiente malarico ed emoglobinopatia ha permesso l’instaurarsi di un polimorfismo bilanciato, in quanto gli eterozigoti asintomatici godono di un vantaggio selettivo nei confronti delle infezioni da malaria (➔ polimorfismo).
S. dei caratteri poligenici
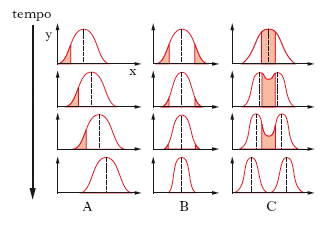
Poiché il meccanismo di azione della s. opera sulla combinazione genotipo/feno;tipo di un organismo, essa agisce anche sui caratteri poligenici o quantitativi (➔ polimeria) che sono controllati da un certo numero di geni e che sono molto suscettibili alle influenze ambientali. Questi caratteri quantitativi, tra cui il peso del corpo, l’altezza, il colore della pelle, mostrano nella popolazione una variazione continua dei fenotipi rappresentata da una curva di distribuzione a campana. Per questi caratteri la s. può essere classificata in direzionale, stabilizzante e disruptiva (o diversificante; v. fig.). La s. direzionale è il meccanismo di s. più importante per gli allevatori di piante e animali che si prefiggono di selezionare caratteri desiderati, spesso rappresentati da fenotipi estremi. Infatti, quando il carattere è poligenico, i fenotipi estremi appariranno nella popolazione solo dopo una prolungata selezione. Un esempio è la s. orientata al contenuto di olio nel chicco del granturco, che in seguito a s. verso l’alto è stato aumentato di tre volte, dal 4% al 15% in circa 50 generazioni, senza raggiungere il plateau. Similmente mediante s. verso il basso si può ottenere una graduale riduzione del contenuto di olio fino all’1%. La s. direzionale favorisce i fenotipi estremi e tende a produrre uniformità genetica nella popolazione. In natura la s. direzionale può avvenire quando, in conseguenza di cambiamenti ambientali, uno dei fenotipi estremi è sottoposto a una s. a favore o contro. La s. stabilizzante invece tende a favorire i tipi intermedi, in quanto vengono selezionati contro ambedue i fenotipi estremi. Uno dei migliori esempi dell’effetto della s. stabilizzante è data dal peso alla nascita in rapporto alla sopravvivenza nel neonato: la mortalità infantile aumenta drammaticamente quanto più ci si allontana al di sotto e al di sopra del valore ottimale di massa corporea alla nascita di 3400 g. A livello genetico una popolazione, mediante la s. stabilizzante, tende al raggiungimento della migliore condizione di adattabilità al suo ambiente. La s. disruptiva agisce contro i fenotipi intermedi e a favore di ambedue i fenotipi estremi e può essere considerata l’opposto della s. stabilizzante. Per es., in una popolazione di insetti predati da uccelli, tutti gli individui di aspetto più vicino al tipo medio sono anche quelli di tipo più frequente e quindi più predati dagli uccelli insettivori, mentre gli individui di aspetto estremo, e quindi più rari nella popolazione, sono meno esposti alla predazione: di conseguenza i fenotipi più comuni sono svantaggiati rispetto a quelli più rari.
S. artificiale
Il processo di s. artificiale si basa sullo stesso meccanismo della s. naturale con la differenza che la pressione selettiva è esercitata dall’uomo sia in maniera inconsapevole, quando fa scelte di cui non ha valutato l’impatto ambientale, sia in maniera consapevole, quando si prefigge di ottenere ibridi o specie di piante e di animali con caratteristiche utili o apprezzabili a livello economico-sociale. L’uso indiscriminato del DDT ha portato a un progressivo aumento della resistenza degli insetti all’azione tossica di questo insetticida, a cui la maggior parte di essi era molto sensibile al momento della sua introduzione. Analogamente l’abuso delle terapie antibiotiche, la cui introduzione ha sconfitto malattie batteriche spesso letali, ha portato alla s. di numerosi ceppi di batteri resistenti a un ampio spettro di antibiotici, costringendo la ricerca farmaceutica alla sintesi di nuove molecole attive.
S. sessuale
La presenza di caratteri sessuali secondari molto vistosi, specialmente in alcuni uccelli e insetti, sembrava in contrasto con i principi della s. naturale, poiché tali ornamenti (per es., le penne del pavone, il brillante piumaggio dei maschi dei fagiani) rendono gli individui che ne sono provvisti più facilmente visibili ai loro predatori naturali. Darwin pensò che per questi caratteri intervenisse un altro tipo di s. che chiamò s. sessuale, basata sul fatto che gli individui forniti di caratteri più vistosi sarebbero più graditi come coniugi e avrebbero perciò maggiore probabilità di riprodursi. Attualmente, riconosciuto il valore fisiologico di molti caratteri sessuali secondari (riconoscimento dei sessi, stimolo all’ovulazione ecc.), si dà una più precisa formulazione alla s. sessuale. In realtà non si tratta di un tipo particolare di s., ma semplicemente di un tipo di s. naturale che opera in uno stadio del ciclo vitale di un individuo, cioè durante il periodo riproduttivo.
Economia
Nell’organizzazione del lavoro, la s. professionale in entrata è una fase del processo di acquisizione di risorse umane che mira a soddisfare le esigenze aziendali di personale. Meno diffusa è la s. professionale in uscita, operata dal lavoratore per confrontare le opportunità offerte dall’impresa e dal mercato esterno, oppure voluta dall’azienda che, nel rispetto della legge, sollecita un’uscita volontaria del lavoratore, solitamente per condizioni negative del mercato.
Nell’economia delle assicurazioni, la s. avversa è un fenomeno originato da asimmetrie informative nella stipulazione di contratti, per cui una variazione delle condizioni contrattuali provoca una s. dei contraenti sfavorevole per la parte che ha modificato, a suo vantaggio, le condizioni (per es., chi ricorre all’assicurazione medica sa della propria salute più dell’assicuratore, e se si considera a basso rischio potrebbe decidere di rinunciare alla copertura assicurativa nel caso di aumento del premio).
Fisica
Regola di s. Ogni regola che limiti la possibilità di una transizione tra stati di sistemi microscopici, come atomi, molecole, nuclei o particelle elementari: poiché nelle transizioni si hanno sempre variazioni di alcuni numeri quantici, le regole di s. si esprimono in genere come variazioni consentite per tali numeri.
Per le regole di s. relative a transizioni elettroniche (➔ spettroscopia).
© Istituto della Enciclopedia Italiana - Riproduzione riservata


