Visione
Dizionario di Medicina (2010)
visione
La visione può essere considerata un processo di interpretazione e di trasformazione di un mondo esterno, fisicamente esistente, nel nostro mondo percettivo mediato dal sistema visivo. Questo processo inizia nella retina, con l’eccitazione di milioni di recettori sensibili alla luce, e ha le sue fasi conclusive probabilmente nella corteccia cerebrale e nelle risposte comportamentali che guidano la nostra interazione con il mondo esterno. Il processo di trasformazione comprende la codifica del segnale visivo in segnali neurali, la loro trasmissione lungo le vie nervose e la loro elaborazione in parallelo nelle diverse stazioni visive, con selezione di quella parte dell’informazione che viene ritenuta più importante, e attenuazione o scarto delle altre, considerate ridondanti. Nella visione è difficile distinguere tra ciò che realmente percepiamo e ciò che invece è semplice inferenza o interpretazione del nostro cervello. [➔ acuità visiva; dispositivi neuromorfi; fototrasduzione; percezione del colore; sensazione e percezione] La v. è una facoltà dovuta all’occhio che, tramite i suoi recettori e altri organi associati, è atto a ricevere come stimolo l’energia raggiante in un certo intervallo di lunghezze d’onda.
La retina dei vertebrati
La distribuzione spaziale di luce (radiazione elettromagnetica di lunghezza d’onda compresa tra 350 e 700 nm) che si forma sulla retina (➔ visione, L’occhio, l’organo della visione) viene trasformata in attività neurale attraverso il processo di trasduzione attuato da fotorecettori, cellule recettrici sensibili alla luce (➔ fototrasduzione). Esistono due tipi di fotorecettori: i bastoncelli e i coni. La retina umana contiene circa 120 milioni di bastoncelli e 6 milioni di coni. Una piccola area della retina, denominata fovea, contiene solo coni. All’interno dei fotorecettori sono racchiuse molecole di fotopigmento, una proteina coniugata composta da due parti legate covalentemente tra loro: l’opsina, la parte propriamente proteica, e il retinale, la parte prostetica, costituita da un lipide sintetizzato dal retinolo (ossia dalla vitamina A). L’opsina può presentarsi in diverse forme: nei bastoncelli si presenta in un’unica forma e il fotopigmento prende il nome di rodopsina (così chiamata a causa del suo colore rosso), mentre nei coni sono presenti tre diverse varietà di opsina, con spettri di assorbimento specifici rispettivamente per i domini del rosso, del verde e del blu. Il sistema visivo è in grado di operare con estrema sensibilità su circa 11 unità logaritmiche di intensità luminose, essendo capace in condizioni ottimali di percepire uno stimolo di una decina di fotoni circa. I coni sono responsabili della visione diurna (fotopica, con luminanza superiore a circa 10 cd/m2), mentre i bastoncelli consentono la visione notturna (scotopica, inferiore a 1 cd/m2). All’interno di ciascuno di questi intervalli di luce ambiente, il singolo fotorecettore deve essere in grado di rilevare variazioni di luce dell’ordine di 6 o più unità logaritmiche. Se l’illuminazione ambientale varia, la sensibilità del fotorecettore varia automaticamente con l’intervallo di sensibilità centrato sul valore di luminanza media (adattamento alla luce). Questo permette di codificare piccoli incrementi di luce che sono proporzionali alla luminanza media ambientale o alla luce dello sfondo, secondo la legge di Weber (➔ sensazione e percezione). Il processo di adattamento alla luce è alla base della costanza della brillanza: una superficie, rispetto al suo sfondo, viene percepita della stessa brillanza indipendentemente dalla quantità di luce che effettivamente riflette. L’adattamento al buio è invece legato al tempo di nuova sintesi del fotopigmento nella sua forma originale, tempo che è molto lento (dell’ordine di minuti) ed è diverso per coni e bastoncelli (➔ fototrasduzione). Proiezioni dei coni e dei bastoncelli. I coni e i bastoncelli differiscono sostanzialmente anche nelle loro connessioni con le altre cellule della retina. Nella fovea, dove sono presenti solo coni, le connessioni recettori-cellule bipolari-cellule gangliari presentano scarsa o nessuna integrazione, fino al caso limite in cui un cono diverge su due cellule bipolari e queste indipendentemente proiettano su due cellule gangliari. Il grado d’integrazione aumenta andando verso la periferia della retina. Per quanto riguarda i bastoncelli, invece, centinaia di recettori convergono su una cellula bipolare e decine di bipolari su una gangliare. È questo diverso grado di convergenza nell’organizzazione nervosa che specializza la periferia della retina, e quindi la visione bastoncellare, nella rilevazione di quantità di luce estremamente piccole. L’alto grado di convergenza comporta che tale regione presenti elevata eccitabilità, ma bassa acuità visiva (➔), intendendo con ciò la capacità di percepire un oggetto nello spazio come distinto da un altro a esso vicino. La fovea, invece, data l’organizzazione delle sue connessioni, presenta bassa sensibilità, ma elevata acuità visiva; essa è anche specializzata nella visione dei colori avendo i coni opsine diverse. Il 5÷10% è costituito da coni sensibili al blu e forma un anello intorno al bordo della fovea. Il resto è costituito da coni sensibili al rosso e al verde in proporzione di 2:1, variabile da soggetto a soggetto (➔ percezione del colore). I recettori, oltre a collegarsi alle cellule bipolari, sono in connessione tra loro mediante le cellule orizzontali. A livello dello strato dei neuroni bipolari si trovano le cellule amacrine, che stabiliscono contatti tra loro, con le cellule bipolari e con quelle gangliari. Circuiti della retina. La circuiteria retinica effettua una sofisticata analisi dell’informazione captata dai fotorecettori, eliminando tutte le informazioni costanti sia nello spazio sia nel tempo ed effettuando un campionamento accurato delle loro variazioni. I campi recettivi delle cellule gangliari sono a simmetria circolare e organizzati in modo antagonistico, con centri che preferiscono incrementi (On) o decrementi (Off) di luce rispetto allo sfondo. L’effetto antagonistico della periferia fa sì che la risposta delle cellule sia massima per oggetti grandi quanto il centro del campo recettivo e fortemente attenuata per stimoli uniformi o 4÷5 volte più grandi del centro. Le cellule gangliari P (Parvo) ricevono segnali da tipi di coni diversi per il centro e la periferia, mentre le cellule M (Magno) sommano i segnali dei coni M (che hanno il picco di sensibilità per la luce verde) e L (che hanno il picco di sensibilità per la luce rossa), sia al centro sia alla periferia dei loro campi recettivi, e quindi sono deputate solo alla visione acromatica. Le cellule M, che rappresentano circa il 10% di tutte le cellule gangliari, hanno un corpo cellulare relativamente grande e ampia arborizzazione dendritica e quindi un campo recettivo relativamente grande; le cellule P, di gran lunga più numerose, hanno invece un corpo cellulare più piccolo, un campo dendritico meno esteso, e quindi un campo recettivo più ristretto. Le cellule M e P differiscono anche per le proprietà temporali: per es., le cellule di tipo M preferiscono stimoli transienti che alternano velocemente. Le cellule P, invece, hanno principalmente un comportamento tonico e mostrano una risposta ottimale a stimoli costanti nel tempo.
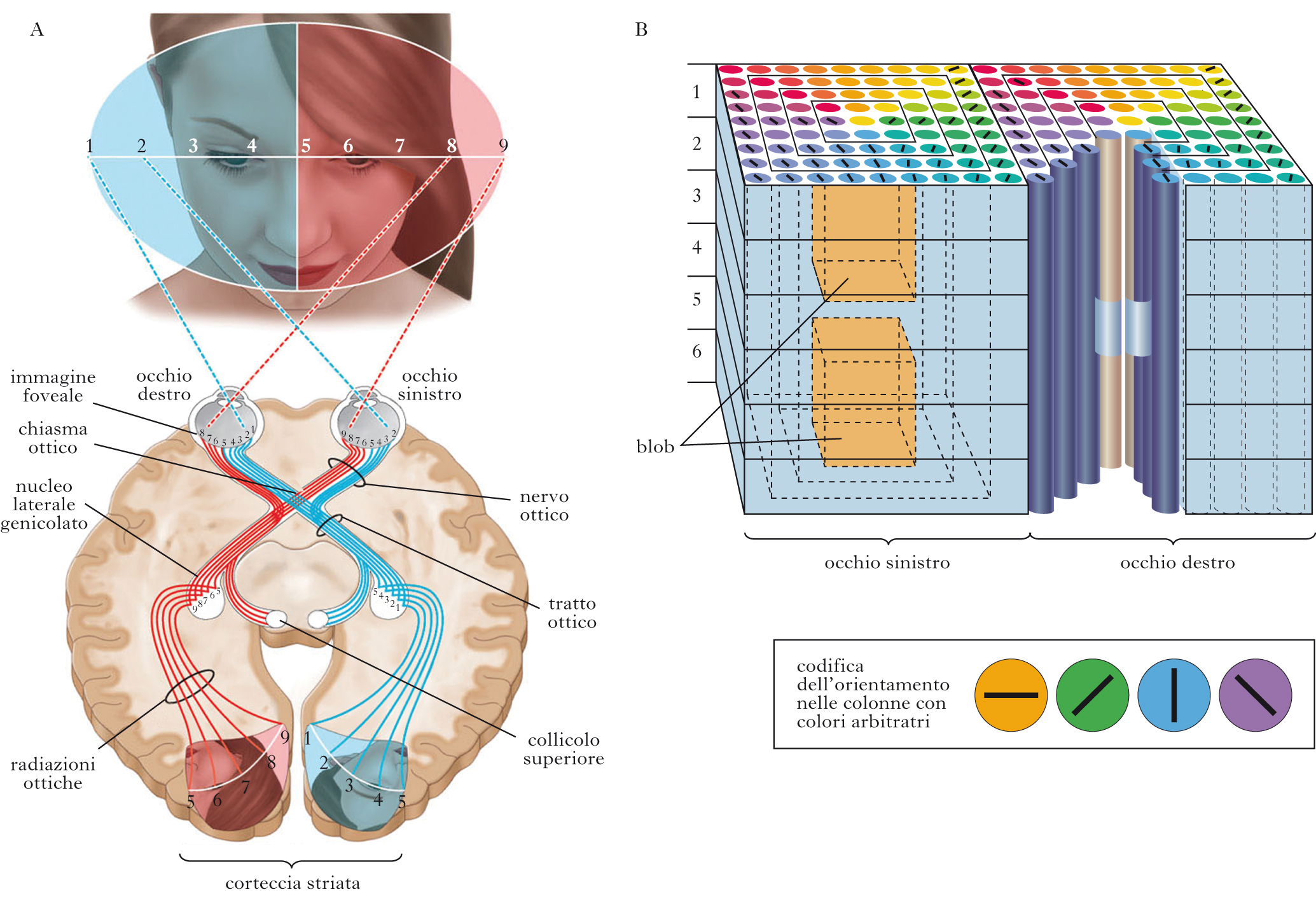
Trasporto dell’informazione visiva dalla retina ai centri superiori
L’organizzazione funzionale della retina porta a una codifica dei segnali luminosi in termini di contrasto sia di luminanza sia cromatico per piccole regioni di spazio e piccoli intervalli temporali. Il contrasto di luminanza è definito come la differenza relativa di luminanza di una regione dello stimolo luminoso rispetto allo sfondo. L’informazione delle cellule gangliari viene trasmessa ai centri superiori, tramite il nervo ottico che è composto da ca. 800.000 fibre. Circa la metà di esse, e precisamente quelle provenienti dalla parte nasale delle due retine, si incrocia nel chiasma ottico e si mette in comunicazione con un nucleo talamico dell’altro lato, il corpo genicolato laterale (LGN). Le fibre provenienti dalla parte temporale delle due retine non si incrociano e si connettono con il corpo genicolato dello stesso lato. Una parte delle fibre del nervo ottico comunica con una formazione nervosa del mesencefalo, il corpo quadrigemino superiore (collicolo superiore). I neuroni dell’LGN proiettano principalmente e direttamente alla corteccia visiva primaria (V1). Dal corpo quadrigemino parte invece una via indiretta che, dopo una stazione nel pulvinar del talamo, raggiunge quasi tutte le aree corticali associative. L’LGN del talamo è una struttura lamellare, con 6 strati. Ogni strato riceve segnali in entrata da un solo occhio: gli strati 2, 3 e 5 ricevono proiezioni dall’occhio ipsilaterale, mentre gli strati 1, 4 e 6 dall’occhio controlaterale. Le cellule negli strati 1 e 2 presentano un corpo cellulare più grande rispetto alle cellule degli altri quattro strati. Per questo motivo, le cellule dei primi due strati sono dette magnocellulari (M) mentre le cellule degli altri quattro sono dette parvocellulari (P). I neuroni degli strati M ricevono segnali in entrata dalle cellule gangliari M mentre i neuroni negli strati P ricevono dalle cellule gangliari di tipo P. A ogni strato P è adiacente uno strato detto koniocellulare, che riceve da cellule gangliari K (cellule bistratificate, opponenti per i coni blu). La corteccia visiva dell’uomo possiede un numero enorme di cellule nervose, dell’ordine di un centinaio di milioni, che operano complesse elaborazioni dei segnali luminosi. Le proiezioni retino-genicolo-corticali hanno una precisa organizzazione topologica: ogni specifica zona della retina proietta a una specifica zona del corpo genicolato e quindi della corteccia. La parte di retina che gode la più ampia rappresentazione corticale è la fovea, in quanto gli input provenienti da questa regione sono i più numerosi. Le differenti parti della retina proiettano a differenti parti della corteccia e del collicolo superiore in maniera ordinata, producendo vere e proprie mappe del mondo visivo, dette retinotopiche.
Corteccia visiva primaria
La corteccia visiva primaria V1 è situata nella porzione occipitale del cervello lungo il solco calcarino ed è detta area striata, o area 17. Lesioni di V1 causano cecità, ma in condizioni di scelta forzata un paziente leso potrebbe mostrare di percepire stimoli visivi senza esserne consapevole. Si suppone che questo tipo di percezione sia mediato da vie subcorticali che oltrepassano V1 e proiettano direttamente ad aree visive superiori. La via M e quella P rimangono in gran parte separate anche a livello della corteccia primaria, mentre le afferenze degli input provenienti dai due occhi sono solo parzialmente segregate, e la maggioranza dei neuroni risponde a entrambi gli occhi. I campi recettivi delle cellule corticali, tanto di tipo M quanto di tipo P, assumono proprietà comples se a opera sia di una convergenza degli input sia di circuiti intracorticali e di feedback che provengono da livelli corticali superiori. A questo grado di complessità i neuroni sviluppano una selettività per proprietà quali: binocularità, orientamento, grandezza, fase spaziale, direzione del movimento e contrasto cromatico. I neuroni con input M e P esibiscono selettività diverse, per es., la via M preferisce stimoli grandi, che si muovono in particolari direzioni e velocemente. L’area 17, così come altre aree associative, presenta un’organizzazione anatomofisiologica partita in moduli (o colonne) che si ripetono periodicamente. Nell’area striata ogni modulo è caratterizzato da neuroni sensibili alle lunghezze d’onda, al movimento e a linee o bordi di particolare orientamento all’interno di una specifica porzione del campo visivo. All’interno di ogni modulo le caratteristiche di risposta delle cellule hanno una disposizione sistematica. La struttura principale è rappresentata dalle colonne di dominanza oculare, che ricoprono circa 1 mm2 di corteccia all’interno della quale sono rappresentati, con pesi variabili, progressivamente e ordinatamente neuroni che ricevono input solo da uno dei due occhi o da entrambi. Interposta a questa struttura colonnare è presente una struttura, detta dei blobs, centrata all’interno della colonna stessa. I blobs contengono cellule a opponenza cromatica semplice o doppia. Ogni blob è posto al centro di strutture radiali che hanno sensibilità per orientamenti progressivamente diversi.
Sensibilità al contrasto e analisi della forma
La sensibilità al contrasto di stimoli modulati sia in luminanza sia in cromaticità è determinata dall’attività all’interno dei moduli, come anche la percezione di caratteristiche elementari dello stimolo, per es. grandezza e orientamento. La sensibilità al contrasto (l’inverso della soglia di contrasto) varia con la grandezza dello stimolo. Se si utilizzano semplici mire sinusoidali, costituite da reticoli, la sensibilità è maggiore per frequenze spaziali intorno a 5÷8 cicli/grado visivo, e si attenua sia per frequenze maggiori (fino ad arrivare ad acuità intorno a 30÷50 cicli/grado), sia per frequenze minori di 1 ciclo/ grado. L’attenuazione alle basse frequenze riflette l’azione inibitoria e antagonista della periferia dei campi recettivi della retina, del genicolato e della corteccia. Il ruolo della selettività corticale per la grandezza e per l’orientamento nel processo percettivo è messo bene in luce da semplici esperimenti che utilizzano tecniche di adattamento. Dopo qualche minuto di esposizione a un reticolo di una certa frequenza spaziale e orientamento, la percezione di un reticolo di frequenza spaziale e orientamento simili, ma non uguali, è fortemente alterata, anche se in modo transiente: l’orientamento e la grandezza delle sue barre vengono percepiti molto diversi da quelli fisici come effetto repulsivo. Inoltre la sensibilità al contrasto peggiora. Questi effetti sono spiegabili ipotizzando che i neuroni corticali di ciascun modulo effettuino una codifica di popolazione per l’orientamento e la grandezza e che l’adattamento generi una riduzione di sensibilità al contrasto dei neuroni che rispondono selettivamente allo stimolo adattante. La simulazione computazionale della codifica dei neuroni per le caratteristiche di base (orientamento, grandezza, fase) con filtri spaziali (lineari o non-lineari) costituisce la base algoritmica di molti sistemi di visione artificiale. Infatti questo è un programma ottimale per la rilevazione di elementi visivi salienti e la successiva segmentazione di superfici contenute nelle immagini visive. A livello dell’area 17 si riscontra il massimo della parcellizzazione dell’informazione visiva. Le aree corticali successive integrano i risultati delle analisi operate dall’area 17 lungo dimensioni specifiche e in parallelo. Tale integrazione si palesa anche nella dimensione dei campi recettivi i quali diventano sempre più grandi a mano a mano che si procede con l’analisi di caratteristiche visive più complesse.
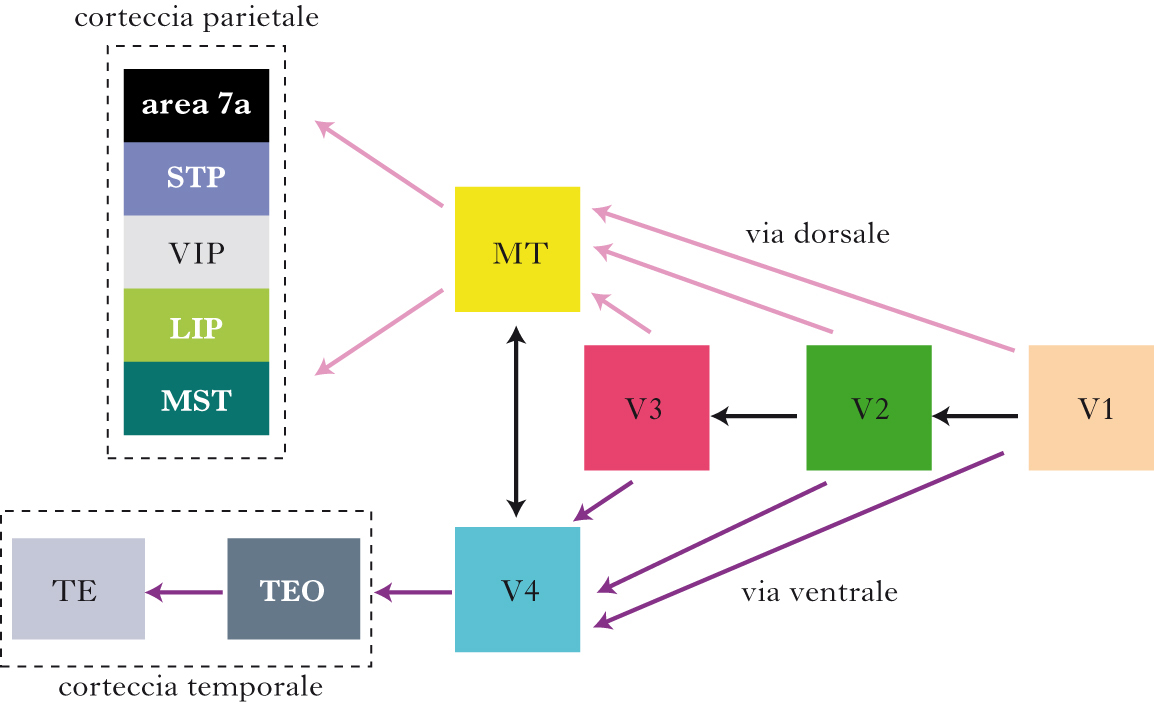
Aree associative visive
Aree associative visive dorsali. Nell’uomo, utilizzando tecniche di imaging funzionale, sono state scoperte più di 40 aree visive che rappresentano l’intero campo visivo su base retinotopica. Molte di queste aree ne hanno una omologa nei primati, mentre altre sono specificamente umane; alcune di esse sono localizzate nell’area 19 di Brodmann. V1 proietta prevalentemente a V2 e l’analisi delle aree associative che ricevono afferenze da V2 può essere schematizzata secondo due ampi sistemi: il sistema what (detto anche via ventrale) – implicato nell’identificazione degli oggetti – e il sistema del where/how (detto anche via dorsale) – coinvolto nella valutazione della posizione spaziale degli oggetti e nell’analisi sensorimotoria alla base dell’azione. Le aree V3, V3A, V3B, V7, V6, V6A, V5 (o MT, Middle Temporal), MST (Medial Superior Temporal), FST (Fundus of the Superior Temporal) e le aree intraparietali come LIP (Lateral IntraParietal), MIP (Medial IntraParietal), AIP (Anterior IntraParietal) e VIP (Ventral IntraParietal) formano la via dorsale, che riceve input prevalentemente dalla via M. Le cellule di V3A, V3B e V7 sono selettive per l’orientamento e si pensa che possano avere un ruolo nell’elaborazione delle forme dinamiche. L’area V5 (o MT) elabora informazioni relative al movimento locale e alla profondità stereoscopica ed è caratterizzata da un’organizzazione colonnare per la direzione del movimento. La stimolazione locale di cellule dell’area MT induce una propensione a percepire il movimento lungo la direzione codificata dalla colonna eccitata, mentre una lesione della medesima porta alla cecità selettiva del movimento. MT proietta a una moltitudine di aree, dette MST, che effettuano analisi di movimento globalmente su tutto il campo visivo. Queste analizzano il flusso ottico, un’informazione successivamente utilizzata dai centri motori superiori, per controllare sia i movimenti oculari sia il movimento del corpo. La via M proietta anche alla corteccia parietale che media l’integrazione del movimento e della profondità nella rappresentazione dello spazio e nei movimenti di grasping (afferramento) e genera una mappa attentiva dell’informazione visiva. La lesione di questa regione negli esseri umani causa una condizione detta sindrome di Balint, caratterizzata da tre sintomi principali: atassia ottica, aprassia oculare e simultaneoagnosia.
Aree associative visive ventrali. La via ventrale è costituita da una moltitudine di aree che ricevono da V2 e V3, come il complesso LO (Lateral Occipital), l’area V4 (la cosiddetta area del colore), e infine dalla corteccia inferotemporale (IT). Queste aree ricevono prevalentemente input dalla via P che alimenta schematicamente due sottosistemi, uno più specializzato nella visione delle forme (sistema interblob di V1) e uno in quella dei colori. L’area IT occupa un’ampia porzione della corteccia dei primati, all’incirca tra il 17 e il 18% della neocorteccia; in essa convergono informazioni sia visive sia mnestiche. In LO le cellule sono attivate da stimoli relativamente semplici, quali strisce e cerchi e, in generale, oggetti, mentre i neuroni della corteccia inferotemporale, i cui campi recettivi sono molto ampi (arrivando a includere tutto il campo visivo), sono attivati da stimoli più complessi. Nell’uomo si osservano due sottoaree IT, una paraippocampale che codifica l’informazione relativa ai luoghi, l’altra disposta lungo il solco fusiforme che codifica i volti. Tutti i neuroni dell’area IT rispondono indipendentemente dalle dimensioni dell’oggetto, dal suo orientamento e dalla sua posizione nel campo visivo: ciò significa che queste cellule sono coinvolte nella percezione visiva dell’oggetto nel suo complesso. La corteccia IT è coinvolta nella percezione del ‘prototipo’ dell’oggetto, sebbene la sua selettività possa cambiare e quindi svolgere anche un ruolo di ‘sistema esperto’ a supporto della classificazione, categorizzazione e memorizzazione degli stimoli visivi. Lesioni bilaterali di alcune zone di questa corteccia producono un disturbo chiamato prosopoagnosia (➔), consistente nell’incapacità, parziale o totale, di riconoscere le facce. Lesioni di altre zone della stessa corteccia, in partic. della sua parte anteriore, causano alterazioni nella visione delle forme o dell’orientamento, senza danneggiare altre funzioni, come l’acuità visiva o la percezione del movimento e del colore. David Burr, Maria Concetta Morrone
L’occhio, l’organo della visione
L’occhio è l’organo di senso per la messa a fuoco dei segnali luminosi e la loro trasduzione in segnali nervosi. I meccanismi ottici utilizzati per la messa a fuoco e la rilevazione degli stimoli luminosi variano in complessità da specie a specie, dalle semplici proteine fotosensibili degli organismi unicellulari agli occhi compositi con guide d’onda degli insetti, agli occhi che utilizzano specchi concavi dei molluschi bivalvi, invece di lenti come nei vertebrati. Molte di queste soluzioni evolutive corrispondono a disegni ottimali che massimizzano l’abilita risolutiva e la sensibilità agli stimoli visivi nei diversi habitat.
Anatomia e fisiologia dell’occhio
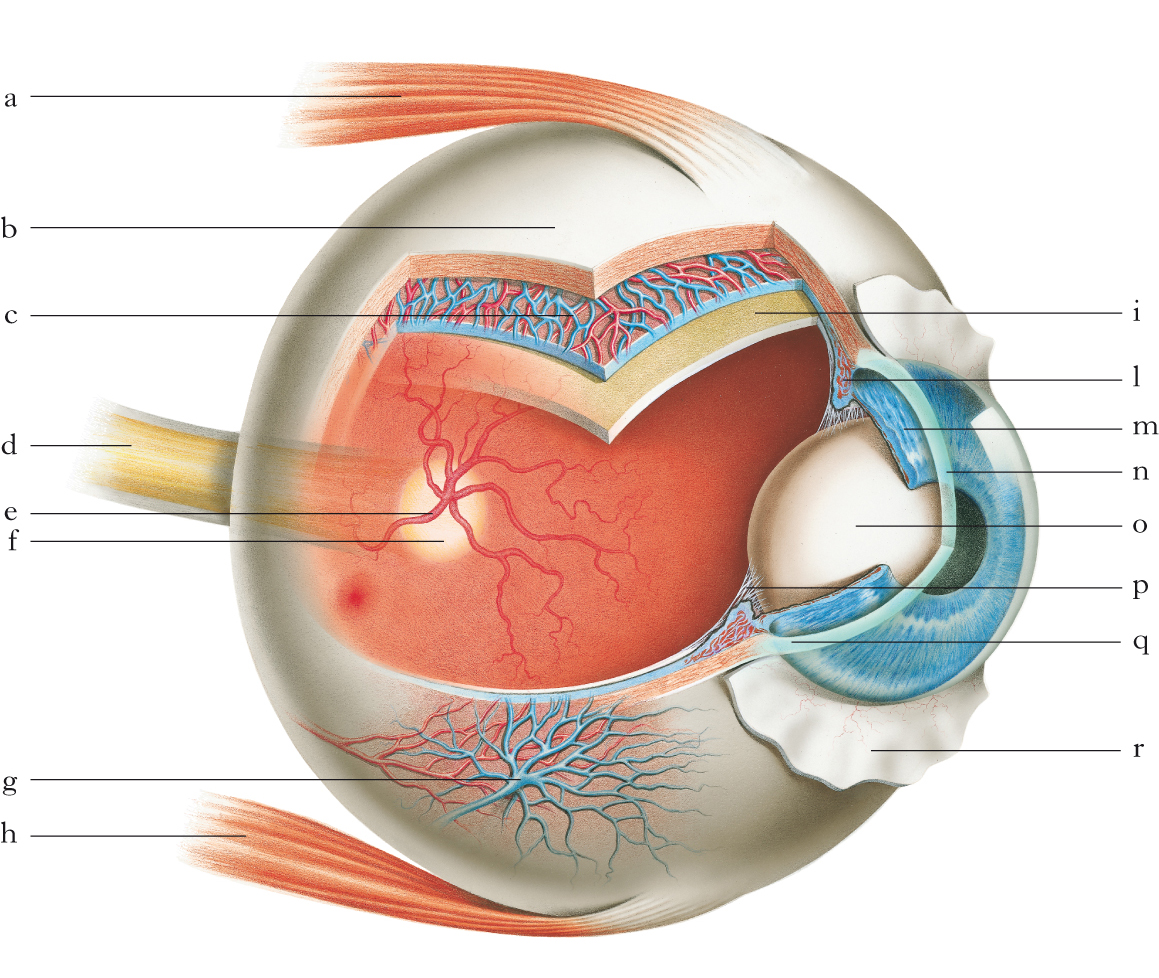
L’occhio dell’uomo ha una forma quasi sferica (bulbo) di circa 24 mm di diametro ed è situato nell’orbita del cranio. La parte anteriore presenta un rigonfiamento anch’esso quasi sferico di circa 12 mm di diametro, costituito da una membrana trasparente detta cornea. Il resto del bulbo è rivestito da una membrana connettivale biancastra, traslucida e anelastica: la sclerotica, o sclera. All’interno della sclera si trova una membrana pigmentata e ricca di vasi, l’uvea, che consta di 3 porzioni: la coroide, con funzione nutritizia grazie alla ricchezza di vasi sanguigni e posta posteriormente; il corpo ciliare, costituito, oltre che da elementi vascolari, da fibre muscolari, il cosiddetto muscolo ciliare, la cui contrazione determina un aumento della curvatura del cristallino e del suo potere diottrico convergente; l’iride, che forma la parte anteriore della tunica vascolare, e precisamente quella che traspare attraverso la cornea e che può assumere diversi colori. L’iride presenta al centro un forame, detto pupilla, che si restringe sino a circa 2 mm sotto l’azione della luce, dilatandosi invece sino a circa 9 mm con l’oscurità (riflesso fotomotore). All’occhio sono annessi la congiuntiva e l’apparato lacrimale che hanno funzioni protettive e ottiche. La tunica nervosa, o retina, tappezza tutta la cavita del bulbo oculare, dal polo posteriore al contorno del foro pupillare; presenta una piccola deflessione nella parte posteriore del quadrante temporale a 5o di eccentricità dall’asse ottico, corrispondente alla fovea. La retina è la membrana fotosensibile che effettua prima la traduzione del segnale luminoso in segnale elettrico e nervoso a opera dei fotorecettori (➔ fototrasduzione), poi un’analisi locale del segnale grazie a circuiti altamente specializzati, inviando le informazioni elaborate ai centri visivi del talamo tramite il nervo ottico che fuoriesce dall’occhio (papilla ottica) a 11o di eccentricità orizzontale nel quadrante nasale della retina. La fovea corrisponde alla porzione di retina con massima densità di fotorecettori. L’occhio ruota per mezzo di sei muscoli extraoculari inseriti sulla sclera. Il globo oculare è protetto da consistenti depositi di grasso che lo circondano all’interno dell’orbita, e da mobili strati di tessuto conosciuti come palpebre. La rapida chiusura delle palpebre (ammiccamento) ha la funzione di detergere e idratare la superficie dell’occhio con il liquido lacrimale che ricopre piccole imperfezioni ottiche. In condizioni normali, automaticamente chiudiamo le palpebre circa una volta ogni 4 s e il ciclo di completa chiusura e riapertura dell’occhio necessita di circa un terzo di secondo: per meta di questo tempo, le palpebre restano completamente chiuse, riducendo del 90% la luce che raggiunge la retina. Raramente percepiamo questi oscuramenti transienti perché selettivamente soppressi centralmente. I mezzi rifrangenti dell’occhio, che i raggi luminosi devono attraversare prima di giungere a stimolare gli elementi sensoriali della retina sono: la cornea, l’umor acqueo, il cristallino (lente biconvessa dietro la pupilla), il corpo vitreo (massa trasparente e gelatinosa). L’ottica dell’occhio corrisponde a una lente variabile da ca. 60 a 72 diottrie (D), di cui circa 44 sono fornite dalla cornea e la restante parte dal cristallino che, variando curvatura, varia il suo potere diottrico da 16 D a 28 D. La funzione del cristallino è quella di mettere a fuoco sulla retina l’immagine di oggetti a diverse distanze dall’occhio (accomodamento). Il cristallino non è omogeneo, ma l’indice di rifrazione varia per i diversi strati di cui è composto crescendo verso l’interno. L’asse visuale dell’occhio corrisponde a una linea che passa dal punto nodale (che si trova circa a 7 mm dal vertice corneale) e dal centro della fovea e corrisponde alla posizione retinica dell’immagine fissata. L’asse visuale non coincide con l’asse ottico del sistema, da cui dista di ca. 5o di angolo visivo. La qualità di un’immagine dipende dalla diffrazione della pupilla, dalle aberrazioni introdotte dalla cornea e dal cristallino. Ulteriori aberrazioni vengono generate da piccoli difetti di curvatura dei mezzi ottici e questi errori si sommano. Tuttavia gli errori della cornea e del cristallino spesso si compensano, cosicché la qualità del sistema risulta migliore di quella delle singole componenti. L’immagine di un piccolo punto all’infinito in un occhio con una pupilla di 3 mm produce un cerchio con diametro di circa 0,8' d’arco visivo (➔ acuita visiva) dovuto sia alle aberrazioni ottiche sia alla diffrazione della pupilla. Questo valore corrisponde a circa 2 volte lo spessore degli elementi fotosensibili più piccoli della retina. Patologie dell’occhio. Relativamente frequenti sono i disturbi della mobilità oculare, i vizi di rifrazione dell’occhio che comportano un’errata messa a fuoco dell’immagine (miopia, ipermetropia, astigmatismo), le alterazioni di suoi elementi strutturali come cornea (cheratiti), uvea (uveiti), retina (retinopatie, distacco retinico), cristallino (cataratta); i processi infiammatori a carico degli annessi oculari come congiuntiva (congiuntivite), palpebre (blefariti), sacchi lacrimali (dacriocistite), ecc.


