Memoria immunologica
Enciclopedia della Scienza e della Tecnica (2007)
Memoria immunologica
È noto ormai da molto tempo che la prima esposizione a un agente infettivo determina spesso in un organismo la capacità di salvaguardarsi per tutta la vita da un'infezione successiva. Oggi è comunemente accettata l'opinione che la resistenza alla seconda infezione rifletta una condizione durevole di memoria immunologica contenuta nei linfociti T e B. Le risposte all'antigene di tipo 'memorizzato' si verificano tipicamente in modo più rapido e più intenso rispetto alla risposta primaria, conducendo alla pronta eliminazione dell'agente patogeno coinvolto. Il ritmo accelerato delle risposte memorizzate va attribuito soprattutto all'aumento della frequenza delle cellule che maturano diventando dei linfociti capaci di rispondere all'antigene. Inoltre, la sensibilità all'antigene delle cellule della memoria risulta acuita dalle loro particolari caratteristiche fenotipiche, che le differenziano dalle cellule vergini.
Allo scopo di comprendere le proprietà e il destino delle cellule della memoria è importante prendere in considerazione, in primo luogo, le caratteristiche dei linfociti vergini e il destino delle cellule che partecipano alla risposta primaria. La risposta immunitaria primaria agli agenti infettivi determina una marcata espansione delle popolazioni di cellule T e B specifiche per l'antigene e la loro differenziazione in cellule effettrici, quelle che colpiscono e cercano di eliminare l'antigene. È il cosiddetto 'processo di selezione clonale', che rende generalmente molto specifica la reazione dell'organismo agli agenti patogeni. La maggior parte dei linfociti reattivi viene eliminata al termine della risposta primaria, ma alcune cellule sopravvivono e si differenziano in cellule della memoria di lunga vita. La memoria immunologica costituisce la base per la risposta immunitaria secondaria, che interviene in caso di successivo contatto con un agente patogeno già incontrato in passato. Questa risposta secondaria (ben noto è l'esempio della varicella) risulta in genere più rapida ed efficace di quella primaria, proprio grazie all'esistenza di queste cellule longeve.
La maggiore efficienza della risposta secondaria riflette una crescita della frequenza delle cellule che danno vita alle cellule effettrici. Inoltre, le cellule originate dalle cellule memoria esibiscono una maggiore affinità per l'antigene rispetto alle cellule linfocitarie che sono rimaste dalla prima infezione. I complessi fattori che governano la generazione e la sopravvivenza delle cellule memoria del sistema immunitario non sono stati ancora del tutto chiariti. Il fenomeno della memoria è probabilmente dovuto alla sovrapposizione di più cause. È possibile che il sistema immunitario venga continuamente stimolato per mezzo della presenza dell'antigene: è noto infatti che, per alcune malattie, gli antigeni rimangono presenti nell'organismo anche molto tempo dopo la risposta primaria all'infezione. Sembra comunque che sia più importante la stimolazione dell'immunità grazie alla continua produzione di citochine (come l'interferone di tipo 1 e l'interferone γ). La continua stimolazione mantiene le cellule memoria in uno stato di semiattivazione: ciò renderebbe possibile la rapidità caratteristica della risposta secondaria.
Risposta immunitaria primaria e destino delle cellule effettrici
La maggioranza delle cellule T e B di animali giovani sono immunologicamente vergini. Esse sono generate negli organi linfatici primari ‒ nel timo le cellule T e nel midollo osseo le cellule B ‒ dai quali migrano agli organi linfatici secondari, cioè la milza, i linfonodi e le placche di Peyer. In queste sedi le cellule T e B non si trovano sotto forma sessile, bensì sotto forma di una popolazione di linfociti circolanti, che migrano in continuazione da un organo all'altro tramite il sangue e la linfa. La durata massima della vita delle cellule T e B vergini è da lungo tempo controversa: benché molti immunologi le considerino come cellule intrinsecamente di vita breve, avviate a morire entro poche settimane in mancanza dell'esposizione all'antigene specifico, oggi risulta accertato in modelli animali che i tipici linfociti T e B con fenotipo vergine mostrano caratteristiche che indicano un ricambio cellulare lento. Nell'uomo, gli studi condotti hanno indicato che le cellule T con fenotipo vergine possono rimanere in questa fase intermedia per un periodo di molti anni. Benché l'invecchiamento sia associato a un declino graduale della proporzione di cellule con fenotipo vergine, queste ultime non scompaiono del tutto, sono anzi ancora visibili in età molto avanzata. La generazione ex novo di linfociti vergini, molto pronunciata prima della pubertà, potrebbe continuare, a un tasso ridotto, per tutta la vita. Dunque, benché ciò non sia confortato da prove dirette, non è escluso che alcune delle cellule con fenotipo vergine formatesi in età avanzata siano cellule di nuova formazione.
Attraverso quella che definiamo 'ricircolazione linfocitaria', le cellule reattive all'antigene sono mobilitate in tutto il corpo, giungendo ad accumularsi nelle stazioni in cui si concentra l'antigene, precisamente la milza e i linfonodi drenanti. In queste sedi, gli antigeni vengono inglobati da cellule specializzate per la presentazione dell'antigene (APC, Antigen presenting cells), quali i macrofagi e le cellule dendritiche, e poi esposti sulla loro superficie sotto forma di peptidi legati a molecole MHC (Major histocompatibility complex) di classe I e di classe II. L'incontro con l'antigene presentato dalle APC comporta l'allontanamento delle cellule T dalla popolazione di linfociti circolanti e il loro temporaneo isolamento, con il quale ha inizio la risposta immunitaria. Questo stesso fenomeno si verifica anche per le cellule B, sebbene il loro isolamento sembri scaturire dal contatto con l'antigene libero piuttosto che con l'antigene processato dalle APC.
Il contatto con l'antigene provoca la proliferazione delle cellule T e B specifiche per l'antigene e il loro differenziamento in cellule effettrici. Queste ultime lasciano gli organi linfatici dopo vari giorni, diffondendosi in tutto il corpo attraverso il flusso sanguigno. Dopo l'eliminazione degli agenti patogeni, le cellule effettrici risultano in soprannumero; la maggior parte di esse muore rapidamente. È presumibile che il deperimento di un gran numero di cellule effettrici al termine della risposta immunitaria rifletta l'esigenza di conservare il repertorio primario di linfociti vergini; se alle cellule effettrici fosse infatti consentito di sopravvivere indefinitamente, il sistema linfatico diverrebbe presto dominato da un ristretto repertorio di cellule T sensibilizzate, incapaci di rispondere all'aggressione di nuovi agenti patogeni. La distruzione delle cellule effettrici che segue la risposta immunitaria evita proprio che si presenti questo problema.
Generazione e fenotipo di superficie delle cellule della memoria
Il fatto che la maggior parte delle cellule effettrici abbia vita breve implica che le cellule della memoria possiedano proprietà particolari, in virtù delle quali possono resistere ai meccanismi di apoptosi. A questo proposito, viene spontaneo porsi una domanda, alla quale non si è ancora trovata risposta: le cellule della memoria sono la diretta progenie delle cellule effettrici o piuttosto evolvono da uno stadio precoce della risposta iniziale? Nonostante i numerosi studi sperimentali e le molte ipotesi avanzate, non vi sono ancora dati sufficienti per corroborare l'uno o l'altro modello. Sia per le cellule B sia per le cellule T non si ha una chiara idea della loro origine e della loro relazione con le cellule effettrici generate durante la risposta primaria. Si sa tuttavia che le cellule B della memoria vengono generate in un'unica sede, e precisamente nei centri germinativi, all'interno dei tessuti linfatici.
Fenotipo di superficie delle cellule della memoria
Mentre i linfociti vergini sono cellule in stato di riposo, le cellule della memoria dispongono in genere di un fenotipo attivato o semiattivato e presentano perciò una parziale somiglianza con le cellule effettrici. Per via del loro stato di attivazione, esse sono in genere sostanzialmente più grandi delle cellule vergini, inoltre dispongono di un'ampia gamma di marcatori di attivazione, per esempio i recettori dell'interleuchina 2 (IL-2R), e sono caratterizzate da un alto tasso di proliferazione. Il fenotipo delle cellule della memoria varia nelle diverse fasi che seguono la risposta primaria: se esso viene analizzato entro le prime settimane dalla fase di risposta, si rileva che molte delle cellule della memoria, a causa del continuo contatto con l'antigene a concentrazione via via decrescente, stanno compiendo il loro ciclo cellulare, e sono dunque in uno stato di manifesta attivazione. Anzi, le prime cellule della memoria dispongono solitamente di una funzione effettrice, e ciò implica che la prima memoria sia controllata dalle cellule effettrici residue, generate negli ultimi stadi della risposta immunitaria.
Entro uno o due mesi dall'infezione le cellule T sensibilizzate con manifesta funzione effettrice diminuiscono; il contatto con l'antigene specifico risulta notevolmente ridotto e le cellule T sopravvissute si mutano ‒ seppur in modo incompleto ‒ in cellule con fenotipo in stato di riposo. Come le cellule T, le cellule B che sono coinvolte nella risposta primaria risultano, nella fase culminante, costituite in gran parte da cellule effettrici, ma successivamente, a mano a mano che l'antigene si riduce, si indirizzano verso un fenotipo in stato di riposo. Le tipiche cellule B della memoria presentano, a differenza delle cellule B vergini, una commutazione nell'isotipo dell'immunoglobulina. Un altro tratto distintivo è che esse esprimono solo bassi livelli dell'antigene stabile al calore (HSA, Heat stable antigen), rispetto agli alti livelli espressi dalle cellule B vergini. Allo stesso modo delle cellule T, le cellule B della memoria di lunga vita mostrano un tasso di ricambio relativamente lento, ma esprimono un fenotipo di superficie.
Specificità e funzione delle cellule della memoria
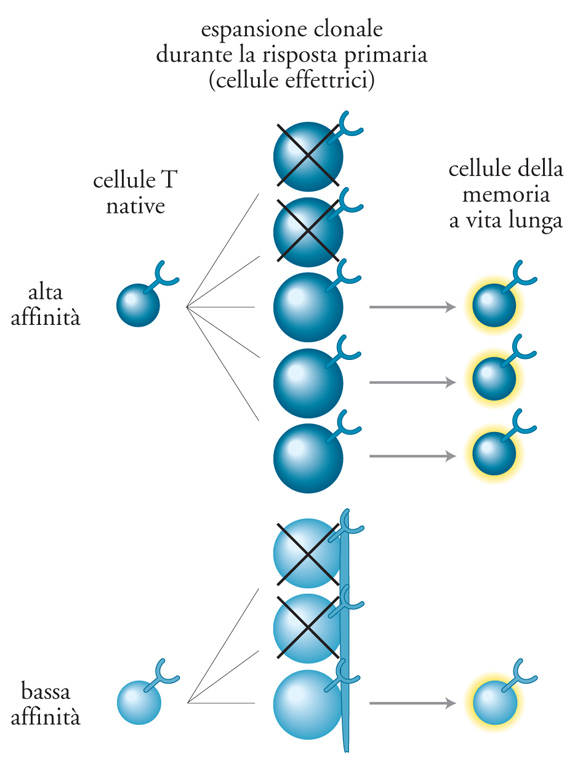
Si è già visto che le cellule T e B della memoria tendono a presentarsi, rispetto ai loro precursori vergini, come cellule con affinità alta, e quindi molto sensibili alla presenza dell'antigene, anche quando questo è presente in basse concentrazioni. Esse sono quindi in grado di scatenare una risposta potente fin dai primi stadi dell'infezione secondaria. Inoltre, la frequenza del precursore delle cellule specifiche per l'antigene è, in genere, sostanzialmente più elevata rispetto a quella osservata nella risposta primaria. Sono proprio gli effetti combinati della maturazione dell'affinità e dell'aumentata frequenza del precursore che rendono la risposta secondaria molto più intensa di quella primaria, conducendo all'efficace eliminazione dell'agente patogeno coinvolto. I tipi di funzione effettrice generati nelle risposte secondarie sono di solito gli stessi della risposta primaria, quantunque il ruolo svolto dalle sottopopolazioni di linfociti che prendono parte alle risposte secondarie abbia una sua rilevanza peculiare. È stato sottolineato che l'efficienza delle risposte secondarie dipende in larga misura dall'immunità umorale. In modo analogo, una produzione protratta di anticorpi specifici si osserva, quasi invariabilmente, anche nel caso di vaccini efficaci. La produzione di anticorpi è particolarmente importante in relazione alle infezioni da batteri extracellulari e da virus citopatici.
Poiché per la produzione di anticorpi occorre la collaborazione delle cellule T, l'immunità umorale è dovuta a un'efficace istruzione sia delle cellule B sia delle cellule T helper. Le condizioni particolari incontrate durante il processo di istruzione delle cellule T sono di grande importanza, poiché possono dettare il tipo di funzione di cui queste cellule disporranno in seguito, a livello di linfociti della memoria. Tra i molti fattori differenti che possono avere effetti rilevanti, vi sono la densità dell'antigene, l'esposizione alle citochine e la gamma di molecole costimolatrici esposte sulle APC. Di particolare importanza è il tipo di citochine incontrato nella fase di istruzione cellulare che, in presenza dell'interleuchina 12 orienta, per esempio, le cellule T verso un fenotipo TH1 (T helper 1). Diversamente, in presenza dell'interleuchina 4 l'istruzione induce la funzione TH2 (T helper 2). L'osservazione che le funzioni delle cellule della memoria possono essere determinate dalle condizioni incontrate nel corso dell'istruzione iniziale ha chiare implicazioni per la creazione di vaccini appropriati. Questa tematica sta riscuotendo grande interesse e probabilmente diverrà centrale nella discussione sulle cellule della memoria.
Fattori che influenzano la sopravvivenza delle cellule della memoria
In passato, le cellule della memoria venivano considerate alla stregua di cellule intrinsecamente di lunga vita, in grado di sopravvivere per tutta la durata della vita dell'ospite senza richiedere ulteriore contatto con l'antigene specifico. Questa interpretazione è stata messa in discussione alla fine degli anni Ottanta del Novecento, quando si osservò che le cellule della memoria sopravvivevano con difficoltà se trapiantate, a meno che non fossero coiniettate con l'antigene specifico. Per spiegare questa scoperta, si è avanzata l'ipotesi che la sopravvivenza delle cellule della memoria in ospiti nei quali l'istruzione è avvenuta molto tempo prima rifletta un contatto persistente con l'antigene residuo. Infatti, è provato che un antigene può sopravvivere per periodi prolungati, dopo la risposta primaria (sotto forma di immunocomplessi), sulle cellule dendritiche follicolari nei centri germinativi. Tali complessi potrebbero comportarsi come serbatoio di antigeni, inducendo una continua stimolazione di basso livello delle cellule della memoria. È stato inoltre dimostrato che la memoria di lungo termine al livello delle cellule T può essere indotta in topi ai quali manchino le cellule B, mentre la sopravvivenza delle cellule T della memoria non sembrerebbe richiedere la presenza degli immunocomplessi. Un'ipotesi alternativa è che l'antigene persista nella forma di complessi stabili peptide-MHC; tale ipotesi sembra però poco verosimile, dal momento che è difficile immaginare complessi peptide-MHC in grado di rimanere stabili nell'uomo per lunghi periodi. Un'ulteriore possibilità è che l'eliminazione degli agenti patogeni al termine della risposta primaria possa essere incompleta, con l'insorgenza di infezione subclinica in sedi isolate. Questa ipotesi mostra la sua validità per determinati virus, come Herpes simplex e forse il virus del morbillo, ma per quanto riguarda altri virus, come quello dell'influenza, l'eliminazione sembra essere completa.
L'idea che la conservazione della memoria richieda un contatto persistente con l'antigene è stata criticata in alcuni studi, nei quali si è dimostrato che le cellule della memoria possono sopravvivere per periodi prolungati, se trapiantate, in apparente assenza di antigeni specifici. In prima approssimazione, queste scoperte sembrerebbero mettere in evidenza che i linfociti istruiti non necessitano di stimolazione cronica per sopravvivere ma, d'altra parte, provano soltanto che non è richiesto un antigene specifico. A questo proposito, alcuni ricercatori hanno suggerito che la sopravvivenza delle cellule della memoria possa riflettere una reazione crociata con vari antigeni ambientali. Il problema sollevato da questa teoria è costituito dal fatto che, a meno che la diversità degli antigeni ambientali non sia estrema ‒ caso che appare improbabile ‒, la sopravvivenza delle cellule della memoria risulterebbe del tutto fortuita, favorendo le cellule T con ampia capacità di reazione crociata.
Una possibile alternativa è che le cellule della memoria richiedano una stimolazione cronica attraverso le citochine piuttosto che attraverso i normali recettori di membrana (TCR). È un'idea che merita considerazione, poiché determinate citochine, vale a dire l'interferone (IFN) di tipo I e l'interferone γ ‒ prodotti in risposta ai virus e ai batteri ‒ sono in grado di stimolare le cellule T in vivo, in apparente assenza di TCR segnalanti. La produzione di IFN sembra favorire la sopravvivenza cellulare durante la risposta primaria. Almeno per quanto riguarda le risposte agli alloantigeni MHC nel topo, l'eliminazione di gran parte della popolazione di cellule T al termine della risposta primaria viene ridotta quando l'antigene è coiniettato con l'IFN di tipo I. Questo effetto ausiliare dell'IFN potrebbe dar conto del motivo per il quale la generazione delle cellule di memoria in risposta ai virus è di solito molto efficiente. La scoperta che la stimolazione delle cellule T tramite gli IFN aumenta la sopravvivenza cellulare suggerisce che il contatto intermittente con gli IFN, rilasciati nel corso di varie infezioni, possa esercitare una stimolazione non specifica sulle preesistenti cellule della memoria, giocando dunque un ruolo importante nella conservazione della longevità della memoria stessa.
È molto probabile che le cellule memoria siano normalmente soggette a una stimolazione intermittente da parte del TCR segnalante (contatto con depositi di antigeni specifici o antigeni ambientali a reazione crociata) o dall'esposizione a citochine, o forse da entrambi questi fattori. La conservazione di una certa frazione delle cellule della memoria in uno stato di attivazione o semiattivazione, oltre a favorire la sopravvivenza cellulare, potrebbe essere importante per assicurare la comparsa delle risposte memorizzate. Se le cellule della memoria si convertissero tutte in cellule in stato di riposo, la generazione delle risposte secondarie sarebbe, infatti, qualitativamente simile a quella delle risposte primarie, e dunque richiederebbe la stessa lunga catena di eventi. Viceversa, la conservazione di una parte delle cellule della memoria in uno stato di attivazione consentirebbe al sistema immunitario di tenere sotto controllo costantemente le vie d'ingresso dell'antigene, per contrastare l'eventuale riproporsi dell'agente patogeno. In questo modo, le cellule attivate o semiattivate si troverebbero nella condizione di poter attaccare l'agente patogeno a livello periferico in uno stadio molto precoce di ricaduta dell'infezione. La produzione continua di anticorpi specifici intensificherebbe tale processo di reazione immunitaria.
Bibliografia
Beverly 1990: Beverly, Peter C., Is T-cell memory maintained by crossreactive stimulation?, "Immunology today", 11, 1990, pp. 203-205.
Di Rosa, Matzinger 1996: Di Rosa, Francesca - Matzinger, Polly, Long-lasting CD8 T cell memory in the absence of CD4 T cells or B cells, "Journal of experimental medicine", 183, 1996, pp. 2153-2163.
Gray 1993: Gray, David, Immunological memory, "Annual reviews of immunology", 11, 1993, pp. 49-77.
McHeyzer-Williams, Davis 1993: McHeyzer-Williams, Michael G. - Davis, Mark M., Antigen-specific development of primary and memory T cells in vivo, "Science", 268, 1993, pp. 106-111.
Schluns, Lefrançois 2003: Schluns, Kimberly S. - Lefrançois, Leo, Cytokine control of memory T-cell development and survival, "Nature reviews. Immunology", 3, 2003, pp. 269-279.
Sprent 2002: Sprent, Jonathan - Judge, Adam D. - Zhang, Xiuren, Cytokines and memory-phenotype CD8+ cells, "Advances in experimental medicine and biology", 512, 2002, pp. 147-153.
Sprent, Surh 2002: Sprent, Jonathan - Surh, Charles D., T cell memory, "Annual reviews of immunology", 20, 2002, pp. 551-579.
Sprent, Tough 1994: Sprent, Jonathan - Tough, David F., Lymphocyte life-span and memory, "Science", 265, 1994, pp. 1395-1400.
Zinkernagel 1996: Zinkeragel, Rolf M. e altri, On immunological memory, "Annual reviews of immunology", 14, 1996, pp. 333-367
© Istituto della Enciclopedia Italiana - Riproduzione riservata


