Neoplasie
Enciclopedia del Novecento I Supplemento (1989)
NEOPLASIE
Oncologia umana
di Francesco Squartini
sommario: 1. Introduzione. 2. Sequenza degli eventi nella cancerogenesi: a) iniziazione e promozione; b) origine monoclonale; c) fasi dell'evoluzione. 3. Modelli di trasformazione. 4. Meccanismi di progressione: a) selezione di cellule preesistenti; b) selezione clonale mutativa; c) modulazione. 5. Preneoplasia: a) caratteristiche fondamentali; b) evoluzione; c) composizione e interazioni; d) considerazioni generali. 6. Neoplasia: a) composizione; b) evoluzione; c) interazioni fra subpopolazioni cellulari. 7. La neoplasia come sistema: a) significato e regolazione dell'eterogeneità; b) cooperazione cellulare e integrazione strutturale; c) metastatizzazione. 8. Conclusioni. □ Bibliografia.
1. Introduzione
‟Di fronte alla impressionante mole di ricerche e di bibliografia sul cancro accumulatasi durante questo secolo, sta ancora la scarsezza dei risultati utili in pratica. Di fronte alla fioritura di ipotesi per interpretare il problema biologico del cancro, sta l'attuale povertà di valide e accettate conclusioni. [...] È difficile immaginare che soluzioni pratiche di portata rivoluzionaria possano essere raggiunte fino a quando il problema dell'accrescimento neoplastico non avrà trovato una interpretazione biologica soddisfacente nel suo insieme. Ed è perciò da ritenersi un progresso la formulazione di concetti nuovi che trascendono l'aspetto anatomo-clinico attingendo al vasto patrimonio di conoscenze della moderna biologia. [...] Tali concetti, e il linguaggio biologico necessario alla decodificazione del problema, cominciano oggi ad affiorare, offrendosi per l'approfondimento e la meditazione".
Queste parole, tratte da un mio articolo precedente (v. neoplasie: Oncologia umana, cap. 15), sintetizzano assai efficacemente la situazione generale delle conoscenze al momento in cui quell'articolo fu scritto, sulla base dei lavori pubblicati fino a tutto il 1975 e a parte del 1976. Da allora, sul piano della prevenzione, cioè dei risultati sociali, e della diagnosi e del trattamento, cioè dei risultati clinici, non si sono verificati progressi sostanziali, a parte quelli inerenti alla razionalizzazione delle conoscenze precedenti. Le attese, o meglio le speranze, sono invece oggi, come sempre, numerose; pare molto promettente, per esempio, l'utilizzazione di anticorpi monoclonali per guidare le sostanze radioattive o i chemioterapici al bersaglio costituito dalle cellule tumorali. Ma di ciò è prematuro parlare, per difetto assoluto di risultati sedimentati, concreti e affidabili. Invece sostanziali sono stati i progressi delle conoscenze nel campo della biologia dei tumori, concernenti la natura e il significato, così complessi, della malattia neoplastica sperimentale e umana.
È dunque pervenuto a maturazione, nel trascorso decennio, ciò che, sul piano dei concetti e del linguaggio biologico necessario alla comprensione del problema, già affiorava alla percezione quando scrissi l'articolo precedente. Oggi, attraverso lo studio dell'eterogeneità delle popolazioni di cellule tumorali e la definizione dei meccanismi che la regolano, attraverso la comprensione ulteriore dei fenomeni della progressione tumorale, attraverso il chiarimento, seppure ancora incompleto, dei contributi genetici ed epigenetici allo sviluppo di una neoplasia, si può illustrare il problema biologico del cancro tracciandone il profilo e fornendone i particolari di dettaglio con una precisione molto maggiore di quella che era consentita dieci anni fa.
Poiché, come si diceva allora, è difficile che si possano escogitare soluzioni pratiche di portata rivoluzionaria finché non si possiederà un'interpretazione biologica soddisfacente del problema dell'accrescimento tumorale, il progresso fatto in questi anni sul versante biologico rappresenta un motivo di speranza per il futuro trattamento clinico delle neoplasie dell'uomo, benché alcuni risultati acquisiti costituiscano motivo di preoccupazione. I nuovi studi hanno infatti chiarito a sufficienza che i tumori maligni, lungi dall'essere solo ammassi di cellule proliferanti anaplastiche, indifferenziate, mal programmate, disorganizzate e perciò fragili, aggredibili, deperibili, facili prede della chemioterapia o della radioterapia, sono, viceversa, popolazioni cellulari organizzate, ecosistemi integrati di cellule variamente specializzate, in grado di interagire, di cooperare fra loro, di programmare una struttura e uno sviluppo, cioè provviste di capacità logiche, quasi fossero creature intelligenti in grado di competere con l'organismo ospite e di sfruttarne i sistemi. I mezzi attualmente a disposizione (chemioterapici, radiazioni, reazioni immunitarie) per distruggere i tumori maligni si rivelano rozzi, primitivi e inefficaci, come dimostrano gli insuccessi che quotidianamente registriamo.
Non potendosi, in una rassegna breve destinata a un vasto pubblico, parlare compiutamente di tutto senza rimanere nel troppo generico e nel superficiale, in questo articolo mi limiterò a illustrare le neoplasie in quanto prodotti dell'organizzazione biologica, ovvero in quanto organismi biologici integrati o ecosistemi. Perciò altri argomenti di alto interesse attuale, come il problema degli oncogeni (v. Bishop, 1982; v. Weinberg, 1982; v. Anders e altri, 1984), non saranno sviluppati in questa sede.
Per molte informazioni di base, che qui non sono ripetute, se non talora in forma estremamente sintetica, si rimanda all'articolo già citato (v. neoplasie: Oncologia umana), particolarmente ai capitoli sulla storia naturale della malattia neoplastica nell'uomo, sui precursori morfologici, sulla progressione tumorale e sulla diffusione metastatica. Qui si tratteranno, nell'ordine, i seguenti argomenti: la sequenza degli eventi nella cancerogenesi, i modelli di trasformazione, i meccanismi di progressione, la preneoplasia, la neoplasia e, infine, la neoplasia come sistema.
I risultati illustrati comprendono un mosaico di dati sperimentali e clinici concorrenti alla soluzione di un unico problema, essendosi chiarito che non esiste alcuna sostanziale differenza, sul piano del comportamento generale, fra neoplasie dell'uomo e degli animali.
2. Sequenza degli eventi nella cancerogenesi
La storia naturale dello sviluppo di una neoplasia, come concordemente indicano i modelli sperimentali meglio conosciuti e le osservazioni sui tumori umani, può essere suddivisa in almeno tre tempi o eventi successivi, che sono l'iniziazione, la promozione e la progressione (v. Pitot, 1982). Di tali eventi il primo è estremamente rapido, il secondo può essere assai lento, mentre il terzo è in genere lentissimo nell'evoluzione iniziale e comprende a sua volta varie fasi, di cui la prima è quella contraddistinta da lesioni preneoplastiche. L'emergenza clinica del tumore coincide di solito con la fase finale della malattia; infatti, fra il momento in cui silenziosamente la malattia neoplastica comincia e quello in cui ci si accorge della sua presenza intercorre un periodo spesso molto lungo, definito periodo di latenza. Da ciò deriva come corollario che la maggior parte della storia naturale di un tumore si svolge al di sotto della soglia clinica e può essere analizzata solo nei modelli sperimentali o ricostruita a posteriori nell'uomo. Parlando dell'inizio di una neoplasia, un problema ulteriore, che merita attenzione, è se la neoplasia abbia origine da una sola cellula trasformata (origine monoclonale) o da più cellule.
a) Iniziazione e promozione
Identificate dapprima nel modello di cancerogenesi cutanea del topo (v. Berenblum e Shubik, 1947), le fasi dell'iniziazione e della promozione sono state poi confermate in molti altri modelli di cancerogenesi di vari organi (mammella, fegato, vescica, colon, stomaco, polmone, tiroide) e controllate in vitro (v. Pitot e Sirica, 1980). In tutti i casi i caratteri dell'iniziazione e della promozione sono sostanzialmente gli stessi. L'iniziazione è un processo istantaneo e irreversibile dovuto a modificazioni indotte direttamente dal cancerogeno nel DNA cellulare. La promozione è invece un fenomeno lento, ripetitivo e reversibile che può essere influenzato o modulato da numerosi fattori ambientali (v. Bontwell, 1974). Vi sono tuttavia cancerogeni definiti completi, perché in grado sia di iniziare che di promuovere lo sviluppo di un tumore. Altri cancerogeni sono detti incompleti, perché in grado di iniziare le cellule di un determinato tessuto, ma non di portare avanti lo sviluppo del tumore. Al contrario gli agenti promoventi non hanno per definizione la capacità di iniziare le cellule neoplastiche, ma spesso capita che essi promuovano verso lo sviluppo tumorale le cellule iniziate da blandi fattori cancerogeni ambientali, fornendo l'impressione erronea di comportarsi come cancerogeni deboli completi. Molti tumori umani si sviluppano probabilmente in questo modo. Solo accurate indagini sperimentali possono discriminare gli agenti promoventi dai cancerogeni deboli completi (v. Pitot, The natural history..., 1977). Enunciate le regole bisogna tuttavia indicare le eccezioni, le quali richiedono aggiustamenti ed estensioni dei concetti definiti sopra (v. Pitot, The stability of..., 1977). In certe circostanze, documentate da esperimenti limite, il processo di iniziazione può rivelarsi infatti anch'esso modulabile (v. Silagi, 1971) o reversibile (v. Mintz e Illmansee, 1975).
b) Origine monoclonale
I risultati di numerose indagini indicano che i tumori originano dall'espansione clonale di una singola cellula trasformata, in quanto la proliferazione iniziale è monoclonale (v. Fialkow, 1972; v. Barrett e Ts'o, 1978; v. Iannaccone e altri, 1978; v. Tanouka e Tanaka, 1982). Tali indagini sono state eseguite prevalentemente sfruttando il polimorfismo della glucoso-6-fosfato deidrogenasi, i cui due isomeri sono determinati da geni strutturali, ubicati nel cromosoma X. Nelle femmine eterozigoti, poiché un cromosoma X è soppresso, vi è mosaicismo dei tessuti normali per le due forme dell'enzima, in quanto circa la metà delle cellule presentano l'enzima in una forma isomerica e la metà rimanente nell'altra. Nel caso di una proliferazione monoclonale è logico aspettarsi una popolazione di cellule omogenee, cioè con l'enzima nella stessa forma isomerica. I risultati dei suddetti studi hanno condotto a concludere che la maggior parte dei tumori umani sono di origine monoclonale (v. Fialkow, 1976). Ma non mancano dati contrari al riguardo (v. Reddy e Fialkow, 1980; v. Rubin, 1980), che suggeriscono invece la possibilità di un'origine policlonale dei tumori, cioè da più cellule (v. Woodruff e altri, 1982). Prevale ancora tuttavia l'opinione di un'origine monoclonale, seppure in disaccordo con le osservazioni morfologiche sulle fasi precoci dello sviluppo neoplastico, che mostrano proliferazioni spesso multicentriche (v. Willis, 19674), e con l'eterogeneità dimostrata dalle popolazioni di cellule tumorali (v. Fidler, 1978; v. Fidler e altri, 1981).
c) Fasi dell'evoluzione
Dopo l'iniziazione e la promozione, nella storia naturale delle neoplasie si possono seguire varie fasi morfologiche di progressione. La prima è in genere rappresentata dallo sviluppo di lesioni preneoplastiche iperplastiche focali, che solo raramente si palesano a livello clinico come neoplasie benigne o displasie, mentre spesso si mantengono subcliniche anche per lunghissimo tempo. Tali lesioni non mostrano alcun attributo di malignità attuale, ma l'analisi del loro comportamento biologico per mezzo dei trapianti consente di dimostrarne la natura precancerosa (v. DeOme e altri, 1959). La fase successiva è rappresentata dalla trasformazione maligna che si verifica nel microambiente delle iperplasie focali (v. Squartini, 1979). Per la regola della progressione indipendente dei caratteri tumorali può accadere che la proliferazione maligna si mantenga in situ prima di acquisire la capacità di invadere i tessuti adiacenti. Quando la malignità si è verificata, inizia lo sviluppo di una popolazione cellulare autonoma, che potrà essere apprezzata clinicamente soltanto dopo che avrà superato certe dimensioni.
Quindi, nel corso dello sviluppo di un tumore, si verificano solitamente almeno due fasi morfologiche e biologiche fra loro ben distinte, attraverso le quali la malattia procede per progressione e che apparentemente postulano due successive trasformazioni cellulari: quella da tessuto normale a lesione preneoplastica e quella da lesione preneoplastica a tumore maligno (v. Squartini, Storia naturale del..., 1982). La progressione ovviamente non si arresta quando la malignità morfologica è raggiunta, ma prosegue con modificazioni pressoché continue dei più vari caratteri delle popolazioni tumorali. Non vi è dubbio però che il salto principale è rappresentato dalla trasformazione che coincide con la malignità.
3. Modelli di trasformazione
Che un tumore maligno prenda origine da un tessuto normale, senza cioè lesioni morfologiche apprezzabili preesistenti, mediante una sola trasformazione cellulare è possibile, ma raro. La presenza quasi costante di lesioni che precedono per lungo tempo lo sviluppo dei tumori di ogni sede e la caratterizzazione biologica sperimentale di tali lesioni hanno perciò diffuso il generale convincimento che la cancerogenesi richieda in effetti due trasformazioni successive (da cellula normale a preneoplastica e da cellula preneoplastica a tumorale) e hanno quindi focalizzato l'attenzione sulla seconda di tali trasformazioni (da cellula preneoplastica a tumorale), situata al confine tra innocenza e malignità (v. Medina, i contributi del 1975).
Esaminando più da vicino le possibilità che si celano dietro la sequenza tessuto normale → lesione preneoplastica → tumore maligno, si vede tuttavia che sono quattro i possibili modelli di trasformazione e che solo uno di questi presuppone due trasformazioni successive o, come si dice, una cancerogenesi in due tempi. Infatti le lesioni preneoplastiche focali possono rappresentare una manifestazione abortiva indipendente del cancerogeno, sprovvista di capacità di evoluzione e collaterale allo sviluppo senza intermediari di una neoplasia (v. Farber, Putative precursor lesions ..., 1976). Ciò è stato dimostrato per la cancerogenesi mammaria da dimetilbenzantracene nel ratto, dove il cancerogeno induce nello stesso tempo noduli di iperplasia alveolare apparentemente non suscettibili di ulteriore evoluzione e tumori maligni, senza lesioni intermediarie, dai dotti (v. Sinha e Dao, 1974).
Un'altra possibilità è che le lesioni preneoplastiche rappresentino una miscela eterogenea di cellule normali e tumorali, le prime con capacità di ritardare lo sviluppo delle seconde attraverso l'inibizione da contatto o meccanismi analoghi. Anche tale ipotesi ha il suo supporto sperimentale; infatti, mentre il trapianto dei noduli di iperplasia alveolare della mammella del topo (che rappresentano la più comune lesione preneoplastica in questo modello di cancerogenesi) richiede molti passaggi prima della trasformazione maligna, cioè non riesce ad accelerare la normalmente lenta progressione dei noduli in tumore, il trapianto delle cellule costituenti i noduli, dissociate o isolate con procedimenti meccanici ed enzimatici, conduce allo sviluppo di tumori maligni in brevissimo tempo (v. Medina e altri, 1978). Dunque, vi sono cellule maligne inespresse nei noduli di iperplasia alveolare, per cui, nonostante il passaggio attraverso una fase preneoplastica, la trasformazione iniziale nella cancerogenesi potrebbe essere una sola. In un elegante esperimento basato sul trapianto in topi ibridi di cellule mammarie normali e tumorali con differenti antigeni di istocompatibilità, la composizione cellulare e la progressione dei noduli di iperplasia alveolare sono state studiate nei dettagli, ottenendo conferma all'ipotesi di cui sopra (v. Slemmer, 1974). Infatti i noduli sono risultati composti da mosaici di cellule alveolari tumorali e di cellule mioepiteliali normali e si è potuto dimostrare che lo sviluppo di tumori dai noduli coincide con l'acquisizione, da parte delle cellule tumorali alveolari, della proprietà di sopravvivere e di proliferare indipendentemente dalle cellule mioepiteliali normali associate (v. Mintz e Slemmer, 1969). La malignità coinciderebbe quindi con la perdita del controllo esercitato da cellule normali sulle cellule tumorali trasformate nelle lesioni preneoplastiche (v. Slemmer, 1980).
Una terza possibilità è che le lesioni neoplastiche siano composte da una miscela di cellule normali e iperplastiche e che la trasformazione tumorale avvenga nelle cellule normali. Anche in questo caso una cancerogenesi apparentemente in due tempi (preneoplasia, neoplasia) avverrebbe in realtà con una sola trasformazione delle cellule normali presenti entro le lesioni iperplastiche. A sostegno di tale possibilità si possono recare i dati sperimentali dimostranti che i cancerogeni sono in grado non solo di indurre lesioni preneoplastiche, ma anche di accelerare la loro trasformazione maligna (v. Medina e DeOme, 1968 e 1970). Paradossalmente, si dovrebbe concludere che l'unica ipotesi con scarso supporto di dati sperimentali è proprio quella suggerita dall'osservazione morfologica di una cancerogenesi in due tempi con duplice trasformazione cellulare: da cellula normale a preneoplastica e da cellula preneoplastica a tumorale(v. Squartini, 1983). Non vi è dubbio comunque che la preneoplasia rappresenti una fase importante nella storia naturale dei tumori, con proprie peculiarità strutturali, evolutive e di progressione, che verranno illustrate in seguito.
4. Meccanismi di progressione
I meccanismi di progressione tumorale sono vari e possono essere genetici o epigenetici e realizzarsi in maniera spontanea o venire indotti da pressioni selettive sulla popolazione di cellule neoplastiche esercitate dallo sperimentatore (somministrazione o sottrazione di ormoni, trapianto), dall'ospite (reazione immunitaria) o dalle cure (medicamenti). Essenzialmente i meccanismi di progressione possono essere ricondotti a tre, che vengono illustrati qui di seguito.
a) Selezione di cellule preesistenti
La trasformazione di un tumore solido in tumore ascite, cioè in una popolazione neoplastica capace di svilupparsi come sospensione cellulare nel liquido ascitico entro il cavo peritoneale, avviene per selezione di varianti cellulari adatte preesistenti in scarso numero (v. Klein e Klein, 1955). La conversione dei tumori mammari solidi in ascite, come pure il numero di passaggi necessari per la conversione, sono anzi strettamente correlati alla quantità di cellule varianti adatte preesistenti, poiché una popolazione tumorale in cui è presente il 5% di cellule varianti è convertibile immediatamente, mentre se le varianti sono l'1% sono necessarie per la conversione due generazioni di trapianto, se le varianti sono lo 0,1% i passaggi in trapianto necessari salgono a quattro e, al di sotto di questa massa critica minima di varianti cellulari adatte, il tumore risulta non convertibile (v. Klein e Klein, 1956). Dunque la progressione (in questo caso l'acquisizione da parte del tumore della capacità di svilupparsi in ambiente liquido inusuale) è qui dovuta a varianti cellulari preesistenti che richiedono tuttavia una pressione selettiva (il trapianto in ambiente liquido) per divenire la popolazione dominante.
La trapiantabilità dei tumori è governata da determinanti proteici o antigeni di istocompatibilità espressi a livello della membrana cellulare, di cui il più importante e noto è l'H-2. Il successo di un trapianto tumorale dipende largamente dal fatto che tali antigeni siano identici nel donatore e nel ricevente. Nel corso della progressione tumorale gli antigeni di istocompatibilità possono essere perduti: così hanno avuto origine i tumori con raggio di trapiantabilità universale nella specie, quali ad esempio il sarcoma di Jensen o il tumore ascite di Ehrlich, per perdita degli alleli H-2. I tumori che si sviluppano in topi ibridi, se trapiantati nei ceppi dei genitori (specie ove questi siano immunizzati), sono in genere rigettati a causa dell'allele eterozigote H-2. Se un tumore cresce nel ceppo di uno dei genitori ciò suggerisce la perdita di uno degli alleli H-2 (v. Klein, 1963). Uno studio sistematico dell'argomento ha dimostrato l'occorrenza e la riproducibilità del fenomeno, che può essere spontaneo e casuale o determinato da pressioni selettive. I tumori che sviluppano tale variazione isoantigenica acquisiscono la capacità di crescere in uno dei genitori e non nell'altro (v. Klein e altri, 1960). La variazione consiste in una ‛perdita' di antigeni, è casuale, stabile, persistente nei trapianti successivi e si ritiene dovuta a selezione di varianti cellulari preesistenti entro la popolazione tumorale di origine, prive fin dal principio dell'antigene ‛perduto' e stimolate a crescere fino alla dominanza dalla pressione selettiva esercitata a mezzo del trapianto nel genitore con identità isoantigenica (v. Klein, 1963).
Anche la trasformazione maligna che si verifica nel microambiente delle lesioni preneoplastiche iperplastiche focali può essere dovuta a selezione di cellule maligne preesistenti, inespresse a causa, per esempio, del controllo esercitato su di esse dalle cellule non trasforrnate contigue. Infatti il trapianto dei noduli di iperplasia alveolare della mammella del topo richiede moltissimi passaggi prima della trasformazione maligna, del resto non costante, mentre il trapianto delle cellule costituenti i noduli, dissociate o isolate con procedimenti meccanici ed enzimatici, conduce, come già detto, allo sviluppo di tumori maligni in brevissimo tempo (v. Medina e altri, 1978). Ciò suggerisce che le cellule maligne preesistenti inespresse nei noduli possano essere rivelate dalla dissociazione e dal trapianto.
b) Selezione clonale mutativa
Numerosi dati suggeriscono che una selezione clonale mutativa sia il meccanismo operativo primario nelle varie fasi dello sviluppo neoplastico (v. Prehn, 1976). In genere il momento iniziale della cancerogenesi consiste in una modificazione potenziale o latente in una larga zona di tessuto, spesso visualizzabile per la comparsa di una lesione preneoplastica. Ma i dati suggeriscono che questo è il risultato dell'amplificazione clonale di un'alterazione ereditabile manifestatasi dapprima in una sola cellula. Anche le tappe successive della cancerogenesi possono avvenire attraverso un analogo meccanismo di selezione clonale. Le mutazioni spontanee sono frequenti nelle popolazioni di cellule tumorali, per cui una popolazione cellulare maligna non mostra di solito omogeneità genetica. Ciò comporta una proliferazione competitiva di differenti cloni cellulari dentro la stessa popolazione. Se uno di questi è provvisto di una maggiore resistenza agli stimoli ambientali, o di una più elevata velocità di accrescimento, esso può sopravanzare gli altri. In tal modo l'indipendenza di una popolazione di cellule tumorali può progressivamente aumentare attraverso selezioni clonali successive su base mutativa (v. Klein e Klein, i contributi del 1958; v. Prehn, 1976).
Un esempio visivo di questo fenomeno si ha nella progressione dei tumori mammari gravidico-dipendenti del topo. Tali tumori hanno struttura tubulare organoide a forma di placca composta da una miriade di piccoli dotti con terminazione a dava irradiantisi da un centro comune verso la periferia. Essi crescono in gravidanza e spariscono dopo il parto. La progressione, quando vi si manifesta, appare di solito come un'area focale rotondeggiante di struttura diversa entro la placca. Dopo il parto la placca residua regredisce, ma non così l'area di progressione focale, per cui il corso clinico del tumore è irrevocabilmente modificato, dipendendo ora dal potenziale di accrescimento e dal livello di autonomia e di differenziazione cellulare del focolaio di progressione (v. Foulds, 1956; v. Squartini e Severi, 1962; v. Squartini e altri, 1980 e 1981). La reversibilità della neoplasia gravidico-dipendente, la sua durata limitata nel tempo e la natura focale della progressione suggeriscono che questa abbia origine per selezione clonale mutativa dalle cellule dipendenti.
È noto che le popolazioni tumorali possono mostrare notevole eterogeneità del cariotipo e che le variazioni nel numero dei cromosomi continuano nel corso dell'intera evoluzione della neoplasia configurandosi come un altro carattere indipendente di progressione tumorale (v. Koller, 1960). Le neoplasie sono più comunemente diploidi, ipodiploidi o ipotetraploidi (v. Koller, 1972). Nelle linee tumorali mantenute in trapianto il cariotipo spesso devia verso l'aneuploidia (v. Hauschka, 1961), verosimilmente per selezione clonale di varianti con numero cromosomico modale (v. Makino, 1957). L'evoluzione del cariotipo è continua nelle cellule tumorali e non riconosce un punto terminale (v. Hsu, 1961). L'ampiezza della variabilità cariotipica e l'instabilità del cariotipo consentono un'evoluzione donale continua delle popolazioni di cellule tumorali, con generazioni di varianti di volta in volta più adatte all'accrescimento autonomo, o a specifiche funzioni, o alle mutevoli condizioni ambientali (v. Makino, 1956). In rapporto a ciò si è postulata l'esistenza in ogni tumore di una linea permanente di cellule staminali con cariotipo modale da cui tutte le altre linee cellulari con diverso cariotipo deriverebbero secondo le esigenze, con ampia possibilità di rimodellamento plastico della popolazione tumorale quando ciò è necessario (v. Makino, 1956). La variabilità cariotipica si comporta come un carattere elementare indipendente dai caratteri tumorali (farmacoresistenza, antigenicità, ormonodipendenza), che possono essere o non essere correlati con una particolare costituzione cromosomica, ed è soggetta alle stesse regole di progressione tumorale riportate sopra (v. Koller, 1972).
c) Modulazione
L'acquisizione di resistenza ai farmaci, che frequentemente compare come carattere stabile e irreversibile nelle popolazioni di cellule tumorali, può derivare dalla soppressione delle subpopolazioni farmacosensibili e ciò rappresenta un meccanismo idoneo per la selezione e la dominanza di varianti cellulari farmacoresistenti preesistenti in scarso numero (v. Goldie e Coldman, 1979), o può dipendere da meccanismi genetici di selezione clonale mutativa (v. Albrecht e altri, 1972; v. Lee e altri, 1973). Ma numerosi fatti suggeriscono che anche meccanismi epigenetici possono governare la comparsa di varianti cellulari farmacoresistenti nelle popolazioni tumorali (v. Harris, 1973). I test di fluttuazione hanno dimostrato che varianti cellulari stabili resistenti ai farmaci compaiono spontaneamente e a caso nei tumori (v. Harris, 1973). Seguendo i tassi di variazione riguardanti la resistenza all'azoguanina si e visto che essi non cambiano nelle linee cellulari diploidi, tetraploidi o ottaploidi e questa mancanza di effetto della ploidia suggerisce che la variazione (resistenza al farmaco) origini nelle cellule somatiche per cambiamenti dell'espressione fenotipica (v. Harris, 1974).
Anche nel caso della comparsa spontanea o indotta di varianti cellulari resistenti alla vinblastina solfato e alla citosina arabinoside i dati suggeriscono che la modificazione cellulare possa realizzarsi, senza alterazioni cromosomiche, a seguito di un cambiamento casuale in un nuovo assetto stabile di espressione genica (v. Harris, 1974). I meccanismi biochimici attraverso i quali la cellula neoplastica riesce a non subire o a riparare il danno provocato dal farmaco sono numerosi, diversi e ormai noti (v. Curt e altri, 1984); citiamo, fra i tanti, il ridotto assorbimento del farmaco, la ridotta attivazione del farmaco, l'aumento del catabolismo del farmaco o dei suoi metaboliti, l'aumentata produzione di molecole bersaglio, l'aumento della concentrazione intracellulare di molecole neutralizzanti il farmaco, la ridotta sensibilità dell'enzima bersaglio, lo sviluppo di vie o siti metabolici alternativi nelle cellule neoplastiche trattate, l'aumento della capacità riparativa delle lesioni indotte dal farmaco, l'alterazione del recettore per i trattamenti ormonali, ecc. (v. Stock, 1978). Tali meccanismi evidenziano tutti modificazioni fenotipiche delle cellule farmacoresistenti non dipendenti da mutazioni e neppure da considerarsi necessariamente irreversibili. Spesso si tratta infatti di cambiamenti di funzioni metaboliche quali possono verificarsi nei tumori per modulazione.
Un'evoluzione o revisione concettuale recente e di portata notevole, nel campo delle teorie della progressione tumorale, è appunto l'aver considerato possibile la progressione anche attraverso modificazioni fenotipiche transitorie delle cellule neoplastiche (modulazione) oltre che attraverso modificazioni genotipiche permanenti. In passato infatti una certa antitesi era insita nei termini progressione e modulazione: il primo denotava, per definizione, eventi irreversibili, mentre il secondo era (ed è) riservato a indicare variazioni minori e reversibili. Invece le indagini recenti, rivolte soprattutto alla caratterizzazione del fenotipo metastatico, hanno condotto a prendere in considerazione, fra le possibili ipotesi proposte per spiegare la selezione di cellule adatte alla metastatizzazione nei tumori, accanto a quelle della selezione statistica (casualità) e della selezione genotipica per mutazione, anche quella della selezione fenotipica per modulazione (v. Weiss, 1979).
L'ipotesi che esistano compartimenti metastatici transitori nei tumori, formulata in alternativa a quella secondo cui le subpopolazioni metastatiche sarebbero permanenti, in quanto originate per selezione clonale mutativa (v. Weiss, 1979), trova supporto in alcuni dati sperimentali, da cui risulta che il fenotipo metastatico può essere reversibile. La coltivazione in vitro delle subpopolazioni costitutive di un tumore mammario murino e il periodico saggio delle loro capacità di formare metastasi in vivo, per valutare la stabilità del fenotipo metastatico, hanno dimostrato che si possono osservare quattro eventualità e precisamente: a) cellule metastatizzanti che si mantengono metastatizzanti; b) cellule non metastatizzanti che pure si mantengono non metastatizzanti; c) cellule non metastatizzanti che divengono metastatizzanti, secondo i consueti canoni della progressione tumorale; ma anche d) cellule metastatizzanti che divengono non metastatizzanti (v. Miller e altri, 1983). Dunque la direzione della progressione tumorale non è univoca ed è possibile, in linea di principio, l'esistenza, nelle cellule tumorali, di condizioni transitorie e reversibili (metaboliche, funzionali o d'altra natura), che le rendono temporaneamente più adatte alla diffusione metastatica.
5. Preneoplasia
a) Caratteristiche fondamentali
Le lesioni preneoplastiche dei differenti organi e tessuti, seppure apparentemente diverse le une dalle altre, hanno in comune molte proprietà, che qui vengono schematicamente riepilogate e sottolineate. Il termine ‛preneoplasia' si applica ad alterazioni strutturali dei tessuti, morfologicamente riconoscibili, che compaiono molto tempo prima dei tumori maligni, mostrano una particolare tendenza alla progressione neoplastica e perciò comportano un aumento del rischio di trasformazione carcinomatosa. Tali lesioni rappresentano la prima risposta morfologica dei tessuti all'applicazione del cancerogeno, sono indotte dal cancerogeno, ma rivelate spesso da fattori promoventi. Perciò, sebbene l'iniziazione sia rapida o istantanea, il tempo di latenza è in genere assai lungo. Nel modello di tumorigenesi virale della mammella murina è stato dimostrato che i noduli preneoplastici di iperplasia alveolare compaiono come lesioni focali apprezzabili fra i nove e i dodici mesi di età (v. Squartini e altri, 1983), mentre cellule trasformate mespresse, capaci di dare origine a noduli di iperplasia alveolare se trapiantate dopo dissociazione enzimatica, sono presenti nelle mammelle morfologicamente normali fin dal terzo mese (v. Medina, Tumor progression..., 1975).
Le lesioni preneoplastiche sono essenzialmente lesioni proliferative, qualunque sia la natura del cancerogeno usato e la sede di lesione. Ciò è evidente per le lesioni precoci degli epiteli di rivestimento (polipi, papillomi, iperplasie, displasie), come per quelle degli organi parenchimali, fra cui le lesioni preneoplastiche della mammella (v. Squartini, Lesioni preneoplastiche e..., 1982) e del fegato (v. Farber, Hyperplastic areas, hyperplastic..., 1976) sono quelle studiate con maggior profitto nei modelli sperimentali e che maggiormente hanno contribuito a chiarire significato e limiti del concetto di preneoplasia. Ciò giustifica il frequente riferimento a esse nelle esemplificazioni.
Altra caratteristica di riscontro comune nelle lesioni preneoplastiche è la natura focale della proliferazione e la molteplicità dei focolai di proliferazione. Infatti tali lesioni appaiono in genere come proliferazioni multiple nell'organo colpito. I noduli di iperplasia alveolare della mammella murina sono multipli, come le iperplasie lobulari e dottali atipiche della mammella umana (v. Squartini, Lesioni preneoplastiche sperimentali..., 1981 e 1983), i noduli iperplastici nella cancerogenesi chimica del fegato, i polipi del colon, i papillomi della vescica, ecc. (v. Sporn, 1976). La multifocalità è con ogni probabilità dovuta al fatto che i cancerogeni applicati inducono la trasformazione non in una cellula nè in tutte, ma soltanto in alcune cellule sparse (verosimilmente in rapporto allo stato in cui si trova al momento il tessuto bersaglio) e che da ciascuna delle cellule trasformate può prendere origine, sotto l'azione di adeguati stimoli promoventi, una proliferazione focale. Ciò suggerisce un'origine monoclonale dei singoli foci, che però non è sempre stata dimostrata.
Le cellule delle lesioni preneoplastiche focali proliferative sono strutturalmente organizzate in modo diverso dalle cellule normali nel tessuto maturo. Il modulo di nuova organizzazione strutturale può essere vario: può trattarsi di un nuovo tipo di differenziazione, come nelle lesioni vescicali (v. Hicks e Wakefield, 1976), oppure di una regressione ad assetti cellulari più primitivi o meno maturi, come nel caso della cute o del fegato (v. Farber, 1973), o ancora di un livello maggiore di differenziazione funzionale, come accade per i noduli di iperplasia alveolare della mammella del topo (v. Squartini, 1977). L'interruzione o il disturbo, sia in difetto che in eccesso, della maturazione o della differenziazione sembra perciò essere una caratteristica comune a molte popolazioni di cellule preneoplastiche (v. Farber, Putative precursor lesions..., 1976).
b) Evoluzione
Il destino delle lesioni preneoplastiche focali è variabile e largamente influenzato dalle circostanze ambientali. Tali lesioni possono infatti regredire, o rimanere stazionarie per lunghissimo tempo (come in genere accade), o infine evolvere verso la malignità. Quindi la progressione di una lesione preneoplastica non è evento inevitabile (v. Sporn, 1976). Anzi, i modelli meglio conosciuti suggeriscono che, tenuto conto della molteplicità delle lesioni, la progressione è l'evento meno probabile e richiede in genere manipolazioni sperimentali o stimolazioni che, modificando equilibri preesistenti, inducano le cellule maligne presenti nella lesione a rivelarsi attraverso la proliferazione autonoma. Nelle lesioni preneoplastiche della mammella del topo la progressione è stimolata, come già detto (v. Medina e altri, 1978), dalla dissociazione cellulare e dal trapianto, o da idonei, ripetuti stimoli ormonali (v. Squartini, MuMTV..., 1981).
In conseguenza di quanto detto sopra, il tempo intercorrente tra la comparsa di una lesione preneoplastica e lo sviluppo del carcinoma è spesso molto lungo, di mesi o anni negli animali, di molti anni (15-25) nell'uomo. Ciò è favorevole alla preservazione della specie, anche se non dell'individuo colpito, perché sposta verso l'età avanzata (non più fertile) l'emergenza clinica dei tumori (v. Farber, Putative precursor lesions..., 1976). Le cause di questa lunga durata sono per la maggior parte da chiarire, anche se si tende a ritenere importante la sorveglianza immunologica dell'ospite (v. Burnet, 1970). Qui comunque interessa rilevare la sostanziale tendenza alla stabilità delle lesioni preneoplastiche, che occupano il maggior tempo nell'ambito della storia naturale dei tumori.
In linea con il dato precedente, le lesioni focali preneoplastiche sono sostanzialmente stabili, come dimostra il loro comportamento nelle linee trapianto. I noduli di iperplasia alveolare virus-indotti della mammella del topo, trapiantati serialmente nei cuscinetti adiposi mammari, sono capaci di propagazione indefinita (v. Medina, 1973). Dopo il trapianto i noduli crescono nel cuscinetto adiposo sviluppandovi un albero ghiandolare con iperplasia alveolare. Quando il cuscinetto adiposo è riempito fino ai suoi confini, il che richiede circa 10 settimane, l'accrescimento cessa e ciò si ripete nei trapianti successivi. Lo sviluppo di tumori maligni nelle linee di noduli in trapianto è possibile, ma infrequente (v. Medina, Biologic characteristies of.., 1975). Ciò dipende però anche dalla variante di virus causale, che può produrre noduli a bassa o ad alta capacità di trasformazione tumorale (v. Squartini e Bistocchi, 1977). Questa sostanziale tendenza alla stabilità dello stato preneoplastico è tuttavia almeno in parte controllata da fattori intrinseci locali, che meritano approfondimento per la loro indubbia importanza pratica. Infatti, come già detto, nei noduli mammari murini di iperplasia alveolare, la progressione si identifica con l'affrancamento delle cellule tumorali alveolari dal controllo esercitato su di esse dalle cellule normali (v. Slemmer, 1980). Su questa base si può spiegare la stabilità delle lesioni preneoplastiche, la loro tardiva e incostante trasformazione maligna e la lunga durata della malattia neoplastica. Lo sviluppo tumorale sarebbe infatti praticamente procrastinato a quando una rottura di equilibrio o la riduzione delle cellule normali, come può aversi anche semplicemente per l'atrofia postclimaterica conseguente all'età, consentano alle cellule trasformate di sfuggire al controllo realizzando la neoplasia (v. Nandi, 1978).
c) Composizione e interazioni
Le popolazioni cellulari preneoplastiche non sono omogenee. Esse risultano composte invece da una miscela di cellule eterogenee diverse fra loro. Un solo nodulo mammario murino di iperplasia alveolare, se frammentato e trapiantato nei cuscinetti adiposi di ospiti singenici, dà origine a una varietà di accrescimenti fra loro differenti per velocità di crescita, stato secretivo, aspetto istologico, capacità di trasformazione maligna, dipendenza ormonale, ecc. (v. Medina, 1973). Se le linee trapianto vengono continuate, si osserva che in ciascuna i caratteri si conservano stabili e vengono mantenuti indefinitamente. Dunque, a dispetto della possibile origine monoclonale, la lesione preneoplastica focale è composta da subpopolazioni eterogenee di cellule caratterizzate da comportamenti diversi, stabili, ereditabili, variamente assortiti e selezionabili con le procedure di trapianto (v. Medina, Tumor progression..., 1975 e 1983). A tale riguardo vi è perciò analogia fra le popolazioni cellulari preneoplastiche e quelle neoplastiche. Ciò indica che l'eterogeneità cellulare è molto precoce nel corso della storia naturale della malattia neoplastica.
Le lesioni preneoplastiche focali differiscono dai corrispondenti tessuti normali per molti caratteri, comprendenti: le attività metaboliche ed enzimatiche, la dipendenza ormonale, la sensibilità ai fattori regolatori dell'accrescimento, lo stato secretivo, la velocità di crescita, l'espressione di neoantigeni, l'immunogenicità, ecc. Questi caratteri sono tendenzialmente stabili e assortiti indipendentemente nelle varie lesioni. Così nei noduli mammari di iperplasia alveolare del topo l'ormonoindipendenza non è necessariamente correlata con la capacità di trasformazione maligna (v. Medina, Tumor progression..., 1975). Alcuni caratteri dei noduli sono essenziali per la stessa lesione preneoplastica (ad esempio l'acquisizione di ormonoindipendenza, che consente il mantenimento della differenziazione lobuloalveolare in assenza di prolattina), o per la sua trasformazione tumorale (ad esempio l'acquisizione di indipendenza dai fattori di regolazione locali dell'accrescimento, quali la regolazione spaziale reciproca fra dotti o il controllo da parte delle cellule mioepiteliali), mentre altri sono chiaramente non essenziali (ad esempio lo stato secretivo, la produzione di proteine particolari espresse come antigeni, la velocità di accrescimento, ecc.) (v. DeOme, 1965). Ogni nodulo preneoplastico rappresenta dunque una popolazione di cellule portatrici di diversi caratteri, essenziali e non essenziali, capaci di progressione indipendente e nell'insieme responsabili delle caratteristiche biologiche e morfologiche del nodulo in base alle proporzioni secondo cui rispettivamente intervengono nella sua composizione.
Un punto sul quale non si è fin qui riflettuto abbastanza è la natura organizzata a formare strutture delle lesioni preneoplastiche. Una struttura richiede spesso l'associazione fra cellule diverse per morfologia e specializzazione funzionale. Alla composizione delle terminazioni funzionali di una ghiandola apocrina come la mammella partecipano più tipi di cellule epiteliali: le cellule alveolari secernenti, le cellule mioepiteliali contrattili, le cellule di rivestimento dei dotti escretori, oltre naturalmente alle cellule stromali di sostegno, alle cellule endoteliali dei vasi, ecc. Sorge dunque il problema dell'origine e delle correlazioni fra le varie cellule costituenti una lesione preneoplastica strutturata e in particolare si tratta di appurare se la trasformazione interessa più tipi di cellule o uno solo. Come già si è accennato, uno studio, in topi ibridi, sulla composizione cellulare delle lesioni preneoplastiche e delle fasi precoci dei tumori mammari murini con diversa istoarchitettura, che consente il riconoscimento delle cellule in base all'accettazione o al rigetto dopo trapianto retrogrado nei ceppi dei genitori, ha dimostrato che i noduli di iperplasia alveolare sono composti da un mosaico di cellule normali e tumorali, nella fattispecie da cellule mioepiteliali normali associate a cellule alveolari tumorali, e che le seconde non possono sopravvivere o mantenere la lesione preneoplastica strutturata in assenza delle prime (v. Slemmer, 1974). Ciò suggerisce una stretta relazione e un'integrazione fra cellule normali e trasformate nell'origine e nel mantenimento delle lesioni preneoplastiche.
Strette interazioni si osservano inoltre fra le lesioni preneoplastiche epiteliali e il loro mesenchima o la loro trama vascolare. Nel topo il trapianto di mesenchima mammario fetale in mammelle adulte induce iperplasie nodulari lobuloalveolari dell'albero ghiandolare, che con alta frequenza progrediscono verso il carcinoma se gli animali sono portatori di virus del tumore mammario murino o sono stati trattati con cancerogeni chimici (v. Sakakura, 1983). I noduli di iperplasia alveolare e altre lesioni mammarie preneoplastiche focali elaborano un fattore angiogenico in quantità proporzionale alla loro propensione alla trasformazione maligna, per adattare il letto vascolare alle esigenze circolatorie e nutritive della neoformazione maligna che da esse prenderà sviluppo (v. Gimbrone e Gullino, 1976). Su questa base è stato anche proposto di valutare e quantificare la natura preneoplastica di lesioni iperplastiche focali in rapporto al loro potere angiogenico (v. Brem e altri, 1978). L'angiogenesi e lo stimolo del mesenchima sulle strutture epiteliali trasformate rappresentano dunque ulteriori parametri di progressione e promozione delle lesioni preneoplastiche.
d) Considerazioni generali
Nella storia naturale della malattia neoplastica la fase di preneoplasia occupa un posto rilevante per la sua costanza, per la lunga durata, per la posizione cruciale rispetto alla progressione tumorale e per le possibilità pratiche che lascia intravedere riguardo alla diagnosi e alla terapia precoci delle neoplasie. Oggi è importante diffondere il convincimento che queste lesioni sono parte integrante del processo di sviluppo del cancro (v. Gullino, 1977). Poiché non è ancora certo che si potrà riuscire a controllare la progressione della malattia neoplastica in fase avanzata, è essenziale che sforzi maggiori siano dedicati allo studio della malattia nelle sue fasi precoci. La necessità di nuove ricerche che conducano a qualche forma di controllo del cancro durante la sua fase preneoplastica, la fase di malignità in situ o la fase invasiva iniziale discende anche dalla scarsità di informazioni sulla loro progressione. Tali studi potranno contribuire a definire con maggiore precisione le fasi di preneoplasia, i loro rapporti con la crescita invasiva, il limite fra fase reversibile e irreversibile della malattia, attraverso l'identificazione di più accurati marcatori diagnostici che rendano possibile la selezione degli individui esposti al maggior rischio e il controllo del cancro prima della malignità (v. Antony e altri, 1976).
6. Neoplasia
a) Composizione
Il dato sperimentale di maggior rilievo dell'ultimo decennio, nell'ambito della biologia dei tumori maligni, è l'eterogeneità delle popolazioni cellulari neoplastiche, non come evento occasionale ma come fatto costante e fondamentale nell'evoluzione delle neoplasie. Ciò ha modificato in parte le precedenti impostazioni e ha fornito nuove basi concettuali per interpretare la progressione tumorale. L'eterogeneità costante dei tumori contrasta col dato della loro origine monoclonale. Ma alcuni riscontri sperimentali consentono di spiegare tale contrasto.
L'eterogeneità cellulare delle popolazioni tumorali può dipendere o da una loro origine policlonale o da una generazione costante di varianti dopo l'origine monoclonale (v. Talmadge, 1983). Alcuni dati disponibili suggeriscono la seconda ipotesi come la più probabile. Se si isola un clone da una popolazione di cellule tumorali, lo si pone in coltura e si analizza appena possibile la colonia cellulare derivata, questa appare non omogenea. Ciò suggerisce che durante lo sviluppo del clone fino a formare la massa sufficiente per l'analisi, cellule eterogenee si sono generate spontaneamente nella popolazione (v. Kerbel, 1982). Nelle metastasi tumorali in vivo, che verosimilmente hanno origine monoclonale, perché derivano da una o poche cellule contigue trapiantate a formare una nuova colonia a distanza, l'omogeneità cellulare clonale intralesionale è presente nell'80% dei casi all'inizio (finché il focolaio metastatico è piccolo) e solo nel 30% dei casi al termine (quando il focolaio è completamente sviluppato). In tutti gli altri casi vi è eterogeneità intralesionale (v. Poste e Greig, 1982). Dunque l'origine monoclonale non contrasta con l'eterogeneità tumorale poiché i tumori di origine unicellulare possono generare rapidamente varianti cellulari con fenomeni diversi (v. Fidler e Hart, 1981).
I tumori sono quindi composti da popolazioni di cellule eterogenee con ogni probabilità originatesi per diversificazione nel corso dello sviluppo. Nell'ambito delle popolazioni eterogenee di cellule neoplastiche, il fenotipo più studiato in questi anni è stato quello metastatico, dal quale ci derivano il maggior numero di informazioni, ma molti altri sono stati studiati. Le subpopolazioni di un tumore possono variare fra loro per molti caratteri, quali il numero dei cromosomi, le proprietà antigeniche, l'immunogenicità, la velocità di accrescimento, le attività enzimatiche e le proprietà metaboliche, la presenza di recettori ormonali, la produzione di secrezioni o di pigmento, la farmacosensibilità, la capacità di invasione, l'attitudine alla metastatizzazione, ecc. (v. Weiss, 1979). Ciascuna sottopopolazione di cellule è responsabile di uno, o più contemporaneamente, dei molti caratteri tumorali che sappiamo passibili di progressione indipendente. Alcune subpopolazioni cellulari possono essere generate per variazione genotipica, altre esprimono invece solo diversità fenotipica modulabile e reversibile. Esempi clinici adatti a illustrare l'eterogeneità per un dato carattere delle popolazioni di cellule tumorali sono forniti dai melanomi metastatizzanti, i cui molteplici focolai si differenziano anche visivamente fra loro per una diversa quantità di pigmento melanico prodotto (v. Sugarbaker, 1979), e dai cancri della mammella a cellule non omogenee per la presenza di recettori ormonali (v. Nenci, 1981). Gli anticorpi monoclonali generati contro cellule metastatiche dei carcinomi mammari dimostrano, nelle colorazioni immunoistochimiche, una notevole eterogeneità di comportamento delle cellule tumorali riguardo all'espressione dei rispettivi antigeni (v. Nuti e altri, 1984). La generazione di varianti che conduce all'eterogeneità delle popolazioni di cellule tumorali è quindi un carattere inevitabile e fondamentale dell'accrescimento neoplastico progressivo (v. Nowell, 1976).
b) Evoluzione
Quando nel tumore si è generato un sufficiente numero di varianti subentra una relativa stabilità dei caratteri. Ciò sembra dipendere da un certo equilibrio che si stabilisce fra le varianti cellulari generate, come se il loro numero e la loro quantità impedissero la generazione di nuove varianti e quindi la comparsa di nuovi caratteri tumorali (v. Nowell, 1976). Se doni di fenotipi cellulari metastatici vengono isolati e studiati attraverso passaggi seriali in vitro o in vivo, si osserva un'alta instabilità, con emergenza rapida di subcloni varianti, diversi per genotipo o fenotipo e per comportamento metastatico, mentre, al contrario, le proprietà metastatiche delle cellule parenti non clonate si mantengono stabili nello stesso periodo, quando esposte alle stesse condizioni di coltura (v. Poste e altri, 1981). Inoltre, ciò che è ancora più interessante, la destabilizzazione del fenotipo metastatico individuale clonato non si manifesta se una serie di cloni sono coltivati insieme (v. Poste e altri, 1981). Ciò indica assai chiaramente una stretta relazione diretta fra eterogeneità cellulare, o presenza di subpopolazioni varianti in sufficiente numero, e stabilità dei caratteri tumorali (v. Talmadge e altri, 1981). Ciò suggerisce anche che: a) la presenza di un numero adeguato di varianti nel tumore da un lato ostacola la generazione di nuove varianti (donde la stabilità dei caratteri tumorali), dall'altro è essenziale per la stessa sopravvivenza tumorale; b) in qualche modo le varianti o i subcloni si influenzano fra loro (v. Poste e Greig, 1982).
Quanto riferito sopra dimostra che ogni selezione di subpopolazioni nell'ambito di un tumore produce instabilità fenotipica innescando automaticamente il meccanismo della generazione di nuove varianti. E ciò che accade in vitro donando le cellule tumorali, ma è anche ciò che accade in vivo quando cellule del tumore primario si isolano istituendo un clone metastatico a distanza (v. Fidler e Hart, 1981); subentra in questo caso una instabilità intrinseca che rimette in moto la diversificazione dei fenotipi cellulari con generazione rapida di subpopolazioni fino al raggiungimento di un nuovo equilibrio (v. Poste e Greig, 1982). Nello sviluppo di un tumore in vivo, pressioni selettive sulla popolazione cellulare neoplastica si possono verificare per motivi molteplici inerenti all'ambiente (ad esempio riserve nutritive limitate), all'ospite (ad esempio reazione immunitaria) o alle cure (ad esempio trattamento farmacologico). Quindi la popolazione tumorale può a più riprese attraversare fasi di instabilità che significano generazione di nuove varianti cellulari con nuove caratteristiche, raggiungendo ogni volta un diverso equilibrio (v. Nowell, 1976). In tal modo la selezione naturale o applicata, precipitando l'instabilità, condiziona e determina il numero di sottopopolazioni presenti nel tumore, l'ampiezza della loro diversità fenotipica e, attraverso questi caratteri, la progressione tumorale (v. Poste e Greig, 1982).
In base a tali vedute la progressione tumorale si identifica con la generazione rapida di subpopolazioni clonali con fenotipi diversi e va considerata come un meccanismo o fattore di diversificazione biologica fondamentale per l'accrescimento delle neoplasie (v. Nowell, 1976). La progressione infatti si manifesta attraverso una serie di cambiamenti indipendenti multipli dei caratteri tumorali dovuti a fenotipi cellulari molteplici e diversi. La diversità dei tipi cellulari presenti nei tumori non è sempre dovuta a mutazioni. Anzi, per la maggior parte, essa è forse la conseguenza di modulazioni reversibili del fenotipo, imposte dal microambiente (nutritivo, metabolico, degenerativo, endocrino, ecc.) o da altre cause (v. Fidler, 1978).
Sebbene una concezione dogmatica radicata consideri la progressione come un evento peggiorativo dei caratteri tumorali in ogni caso, è vero invece che l'evoluzione continua dei tumori non sempre porta alla comparsa di cellule più maligne (v. Fidler e altri, 1978). Quindi la direzione della progressione può, sia pure in casi limite, andare in senso opposto a quello che la parola stessa letteralmente suggerisce. Esempi di questo tipo sono la soppressione reversibile della malignità nelle cellule del melanoma (v. Silagi, 1977), la differenziazione secondo linee embriologiche normali di cellule del teratocarcinoma introdotte in embrioni (v. Illmansee e Mintz, 1976), la regressione dello stato neoplastico nelle piante (v. Meins, 1977) e la già citata perdita della capacità prima posseduta di metastatizzare da parte di subpopolazioni cellulari di tumore mammario murino coltivate in vitro (v. Miller e altri, 1983). L'eventualità di una reversione della progressione rimane tuttavia un fatto molto raro e in contrasto con l'osservazione quotidiana, clinica e sperimentale, dell'evoluzione delle neoplasie.
c) Interazioni fra subpopolazioni cellulari
La presenza di subpopolazioni clonali nei tumori ha sollevato la questione delle possibili interazioni fra esse e stimolato indagini in tal senso, le quali, fornendo una risposta positiva, hanno rivelato nuovi aspetti e significati dell'eterogeneità tumorale. L'analisi di 5 subpopolazioni cellulari derivate da un tumore mammario murino ha appunto dimostrato l'esistenza di varie interazioni fra di loro. Si è visto che le subpopolazioni a lento accrescimento ritardano lo sviluppo di quelle ad accrescimento rapido. Questa osservazione nei trapianti in vivo può essere spiegata in termini di reazioni immunitarie dell'ospite, supponendo che le cellule a lento accrescimento siano più immunogeniche e stimolino la produzione di anticorpi che bloccano anche quelle ad accrescimento più rapido. Ma poiché il fenomeno è riproducibile in vitro, si deve supporre che fattori mibitori prodotti da una subpopolazione possano influenzare la crescita delle altre (v. Heppner e altri, 1980). Il fatto, di non rara osservazione clinica, che la presenza del tumore primario limita l'accrescimento delle metastasi rappresenta forse un fenomeno spontaneo analogo, o meglio spiegabile sulla base di interazioni fra cellule (v. Poste e Greig, 1982). Si è visto inoltre che una subpopolazione di cellule tumorali può influenzare la farmacosensibilità di un'altra, forse attraverso un effetto sul metabolismo del farmaco (v. Miller e altri, 1980).
Da questi fatti emerge un'interpretazione nuova dell'eterogeneità tumorale e un nuovo concetto di neoplasia, secondo cui il tumore policlonale è un ecosistema interattivo, il cui comportamento complessivo è diverso dalla somma dei comportamenti delle sue parti costituenti (v. Heppner e altri, 1980; v. Kerbel, 1982). Dunque la neoplasia va intesa come un sistema policlonale coordinato e intelligente. A loro volta le interazioni fra subpopolazioni di cellule cancellano l'ipotesi di una progressione anarchica, afinalistica dei tumori maligni.
7. La neoplasia come sistema
a) Significato e regolazione dell ‛eterogeneità
Una neoplasia intesa come sistema biologico integrato presuppone non solo proprie regole, ma anche significati, finalità, vantaggi che meritano a questo punto brevi considerazioni.
La generazione costante di varianti cellulari nei tumori mostra analogia con la generazione spontanea di mutanti farmacoresistenti nelle colonie batteriche. La fluttuazione e la diversità impediscono che il tumore sia dominato da una sola o da poche popolazioni cellulari. Uno dei significati dell'eterogeneità sembra perciò essere quello di conferire maggiori possibilità di resistenza e opportunità di sopravvivenza al tumore (v. Poste e Greig, 1982). Un altro significato potrebbe essere quello della specializzazione funzionale.
Sulla base dei dati sperimentali è stato proposto uno schema che spiega i meccanismi di regolazione della fluttuazione di subpopolazioni cellulari durante la crescita tumorale (v. Poste e Greig, 1982). Dopo la trasformazione cellulare maligna, ha inizio una proliferazione clonale seguita assai precocemente da un'emergenza di subpopolazioni tumorali varianti, generate a un tasso sostenuto e costante, finché si giunge a una neoplasia eterogenea con determinati caratteri (N1). A questo punto, cioè quando è stato raggiunto un grado sufficiente di eterogeneità, la generazione di nuove varianti si arresta e si ha stabilità dei caratteri tumorali. Se la popolazione cellulare del tumore viene esposta a selezione (S1), come nel caso di una reazione immunitaria o della somministrazione di farmaci, ecc., si osserva una riduzione significativa del numero di subpopolazioni presenti nel tumore, che tende a divenire più omogeneo. Ciò provoca instabilità genetica della popolazione tumorale, per cui le subpopolazioni che sopravvivono generano nuove varianti a un tasso sostenuto e costante finché è restaurata una diversità adeguata. Di nuovo si avrà quindi una neoplasia eterogenea (N2), diversa tuttavia, per subpopolazioni cellulari e per caratteri, da quella precedente (N1). A questo punto si avrà un nuovo declino o arresto nella formazione di varianti e quindi una nuova stabilità o equilibrio della popolazione tumorale fino a quando un'ulteriore selezione (S2) non rimetta in moto il ciclo di restrizione ed espansione della diversità, che condurrà ancora a una neoplasia eterogenea e stabile (N3), diversa dalle precedenti. Tale processo si può ripetere indefinitamente e attraverso di esso avanza la progressione tumorale. I tumori maligni in progressione avanzata possono rappresentare dunque un mosaico straordinario di subpopolazioni cellulari diverse, che dispiegano modelli pressoché illimitati di variazione fenotipica (v. Poste e Greig, 1982).
I vantaggi biologici offerti da un meccanismo di regolazione come quello di cui sopra sono evidenti. Il più importante è la possibilità di resistere meglio agli assalti dell'ospite o delle cure o a condizioni ambientali sfavorevoli, cioè a tutti i possibili tipi di selezione, che avrebbero invece alte probabilità di successo se le subpopolazioni costituenti il tumore fossero solo poche e poco variate. Lo stimolo alla differenziazione di sottopopolazioni, causato da eventi che ne riducono drasticamente il numero, procura dunque al tumore un potente meccanismo per la sopravvivenza e la rigenerazione. Qualora, anche in assenza di stimoli selettivi esterni, il tumore soggiacesse alla dominanza di una popolazione favorita, il meccanismo della diversificazione si rimetterebbe in moto da solo come fattore di autocorrezione, per contrastare il rischio della dominanza di una sola popolazione, che sarebbe mortale (v. Poste e Greig, 1982).
b) Cooperazione cellulare e integrazione strutturale
Accanto a tali vantaggi vi è il dato delle interazioni fra sottopopolazioni di cellule neoplastiche, che possono consentire una cooperazione cellulare specializzata volta al raggiungimento delle finalità del tumore, per esempio attraverso una regolazione della crescita, in rapporto alle disponibilità circolatorie e nutritive, o un'interazione metabolica (v. Heppner e altri, 1980). Tuttavia si conosce ancora molto poco in questo campo e sono perciò necessarie altre ricerche.
La cooperazione cellulare nei tumori maligni può spingersi fino a un'integrazione fra cellule diverse a formare strutture specializzate. La maggior parte dei tumori umani sono infatti composti da cellule coesive riunite a comporre strutture ghiandolari, tubulari, cordonali, nodi solidi con segni di differenziazione unidirezionale, papille, ecc. Indagini sperimentali recenti hanno indicato al riguardo che nei tumori maligni cellule neoplastiche o normali talora pervengono a formare strutture e che cellule neoplastiche di diverso tipo compongono, associandosi, strutture (v. Slemmer, 1980). La cooperazione e l'integrazione fra sottopopolazioni di cellule tumorali si rivelano inoltre importanti per il raggiungimento del grado più elevato di autonomia e di progressione tumorale, che coincide con la diffusione metastatica della neoplasia.
c) Metastatizzazione
È nota la possibilità di influenzare nei modelli sperimentali la diffusione metastatica dei tumori con manipolazioni varie. Tumori mammari murini incapaci di metastatizzare spontaneamente possono dare luogo a metastasi se le loro cellule costituenti vengono dissociate e iniettate come sospensioni in vena. Ciò può essere spiegato ammettendo la presenza nel tumore di cellule prive di capacità di invadere il circolo, ma in grado di produrre l'attecchimento metastatico a distanza. È stato dimostrato tuttavia anche l'opposto. Vi sono infatti tumori solidi capaci di metastatizzare che perdono questa capacità se le loro cellule isolate vengono iniettate come sospensioni in vena. La spiegazione più logica in tal caso è che le cellule tumorali isolate, non aggregate, non organizzate non sono in grado di dare metastasi (v. Slemmer, 1980). La diffusione metastatica può richiedere dunque in talune circostanze l'integrazione o la cooperazione cellulare. Ciò non stupisce, se si pensa all'estrema complessità dell'evento metastatico, il quale presuppone da parte delle cellule neoplastiche: a) l'adesione alla membrana basale vascolare; la laminina, una glicoproteina della membrana basale, media l'attacco delle cellule tumorali al collageno tipo IV della membrana stessa (v. Liotta e altri, 1983); b) l'invasione della membrana basale; sono state identificate proteasi specifiche nelle cellule tumorali, capaci di degradare il collageno tipo IV e V della membrana basale; queste proteasi sono tanto più abbondanti quanto più le cellule tumorali sono metastatizzanti (v. Liotta e altri, 1983); c) la locomozione attraverso il varco aperto dalla proteolisi nella membrana basale e nel collageno circostante; la direzione di tale locomozione può essere influenzata da fattori chemiotattici (v. Kerbel, 1982); d) la circolazione nei vasi; questa espone le cellule agli anticorpi circolanti e richiede che l'eventuale aggressione possa essere superata o volta a proprio vantaggio; e) l'arresto in punti determinati del circolo; questo arresto, dovuto solo in parte a fattori meccanici, in prevalenza avviene sotto il controllo di fattori chemiotattici (v. Kerbel, 1982); f) l'adesione all'endotelio vascolare per ripercorrere a ritroso il cammino precedente, solubilizzando la membrana basale e l'interstizio; ciò richiede attività enzimatiche specifiche nei confronti del collageno, della fibronectina, della laminina, dei proteoglicani; g) l'attecchimento nel nuovo distretto attraverso la stimolazione dell'angiogenesi, che consenta la crescita tumorale e la ricostruzione di strutture (v. Gullino, 1983). Queste, e verosimilmente non solo queste, sono le difficoltà e le prove che le cellule di un tumore maligno debbono superare per poter metastatizzare (v. Poste, 1983). Superare brillantemente una o due prove non è infatti sufficiente a compensare il fallimento nelle altre. Perciò le cellule metastatizzanti dei tumori maligni sono state paragonate a decatieti, i quali debbono superare bene tutte le prove per giungere al risultato (v. Poste e Fidler, 1980). È realistico pensare che singole cellule possano completare tutte le fasi necessarie alla metastatizzazione? Ciò non si può escludere; forse però è più logico supporre che la diffusione metastatica dei tumori richieda una specializzazione funzionale di più tipi di cellule, la loro cooperazione e la loro integrazione.
8. Conclusioni
Da un rapido confronto con il contenuto del mio articolo precedente (v. neoplasie: Oncologia umana) si può valutare quanto i concetti sull'evoluzione naturale dei tumori maligni siano cambiati in questi anni, quanto sia più complesso il problema e quanto lontana ne sia la soluzione. Tuttavia la consapevolezza ormai acquisita che le neoplasie sono non un accidente casuale, ma una delle possibili conseguenze dell'organizzazione biologica (v. Foulds, 1969-1975) è al presente importante per indirizzare opportunamente le nuove indagini, delle quali c'è ancora molto bisogno.
bibliografia
Albrecht, A. M., Biedler, J. L., Hutchison, D. J., Two different species of dihydrofolate reductase in mammalian cells differentially resistant to amethopterin and methasquin, in ‟Cancer research", 1972, XXXII, pp. 1539-1546.
Anders, F., Schartl, M., Barnekow, A., Anders, A., Xiphophorus as an in vivo model for studies on normal and defective control of oncogenes, in ‟Advances in cancer research", 1984, XLII, pp. 191-275.
Antony, P. P., Bontwell, R. K., Farber, E., Friedell, G. H., Gullino, P. M., Lipkin, M., Nettesheim, P., Price, J., Sporn, M. B., Early lesions and the development of epithelial cancer (Symposium). Summary statement, in ‟Cancer research", 1976, XXVI, p. 2706.
Barrett, C. J., Ts'o, P.O.P., Evidence for the progressive nature of neoplastic transformation in vitro, in ‟Proceedings of the National Academy of Sciences, USA", 1978, LXXV, pp. 3761-3765.
Berenblum, I., Shubik, P., A new, quantitative approach to the study of the stages of chemical carcinogenesis in the mouse's skin, in ‟British journal of cancer", 1947, I, pp. 383-391.
Bishop, J. M., Retroviruses and cancer genes, in ‟Advances in cancer research", 1982, XXXVII, pp. 1-32.
Bontwell, R. K., The function and mechanism of promoters of carcinogenesis, in ‟Critical review of toxicology", 1974, II, pp. 419-443.
Brem, S. S., Jensen, H. M., Gullino, P. M., Angiogenesis as a marker of preneoplastic lesions of the human breast, in ‟Cancer", 1978, XLI, pp. 239-242.
Burnet, F. M., Immunological surveillance: an evolutionary approach, in Immunity and tolerance in oncogenesis (a cura di L. Severi), Perugia 1970, pp. 45-61.
Curt, G.A., Clenteninn, N. J., Chabner, B.A., Drug resistance in cancer, in ‟Cancer treatment reports", 1984, LXVIII, pp. 87-98.
DeOme, K.B., Multiple factors in mouse mammary tumorigenesis, in ‟Cancer research", 1965, XXV, pp. 1348-1356.
DeOme, K.B., Faulkin, L. J., Bern, H.A., Blair, P. B., Development of mammry tumors from hyperplastic alveolar nodules transplanted into gland-free mammary fat pads of female C3H mice, in ‟Cancer research", 1959, XIX, pp. 515-520.
Farber, E., Hyperplastic liver nodules, in ‟Methods in cancer research", 1973, VII, pp. 345-375.
Farber, E., Hyperplastic areas, hyperplastic nodules, and hyperbasophilic areas as putative precursor lesions of liver cancer, in ‟Cancer research", 1976, XXXVI, pp. 2532-2535.
Farber, E., Putative precursor lesions: summary and some analytical considerations, in ‟Cancer research", 1976, XXXVI, pp. 2703-2705.
Fialkow, P. J., Use of genetic markers to study cellular origin and development of tumors in human females, in ‟Advances in cancer research", 1972, XV, pp. 191-226.
Fialkow, P. J., Clonal origin of human tumors, in ‟Biochimica et biophysica acta", 1976, CDLVIII, pp. 283-310.
Fidler, I. J., Gersten, D. M., Hart, I.R., The biology of cancer invasion and metastasis, in ‟Advances in cancer research", 1978, XXVIII, pp. 149-250.
Fidler, I. J., Gruys, E., Cifone, M. A., Barnes, Z., Bucana, C., Demonstration of multiple phenotypic diversity in a murine melanoma of recent origin, in ‟Journal of the National Cancer Institute", 1981, LXVII, pp. 947-956.
Fidler, I. J., Hart, I. R., The origin of metastatic heterogeneity in tumors, in ‟European journal of cancer", 1981, XVII, pp. 487-494.
Foulds, L., The histologic analysis of mammary tumors of mice. II. The histology of responsiveness and progression. The origins of tumors, in ‟Journal of the National Cancer Institute", 1956, XVII, pp. 713-753.
Foulds, L., Neoplastic development, 2 voll., New York 1969-1975.
Gimbrone, M. A., Gullino, P. M., Angiogenic capacity of preneoplastic lesions of the murine mammary glands as a marker of neoplastic transformation, in ‟Cancer research", 1976, XXXVI, pp. 2611-2616.
Goldie, J. H., Coldman, A. J., A mathematic model for relating the drug sensitivity of tumors to their spontaneous mutation rate, in ‟Cancer treatment reports", 1979, LXIII, pp. 1727-1737.
Gullino, P. M., Considerations on the preneoplstic lesions of the mammary gland, in ‟American journal of pathology", 1977, LXXXIX, pp. 413-430.
Gullino, P. M., Angiogenesis and neoplastic transformation of the mammary gland, in Understanding breast cancer: clinical and laboratory concepts (a cura di M. A. Rich, J. C. Hager e P. Furmanski), New York 1983, pp. 247-260.
Harris, M., Polyploid series in mammalian cells, in ‟Experimental cell research", 1971, LXVI, pp. 329-336.
Harris, M., Phenotypic expression of drug resistance in hybrid cells, in ‟Journal of the National Cancer Institute", 1973, L, pp. 423-428.
Harris, M., Comparative frequency of dominant and recessive markers for drug resistance in Chinese hamster cells, in ‟Journal of the National Cancer Institute", 1974, LII, pp. 1811-1816.
Hauschka, T.S., The chromosome in ontogeny and oncogeny, in ‟Cancer research", 1961, XXI, pp. 957-969.
Heppner, G. H., Miller, B., Cooper, D. M., Miller, F.R., Growth interactions between mammary tumor cells, in Cell biology of breast cancer (a cura di C. M. McGrath, M.A. Rich e M. J. Brennan), New York 1980, pp. 161-172.
Hicks, R. M., Wakefield, J. St. J., Membrane changes during urothelial hyperplasia and neoplasia, in ‟Cancer research", 1976, XXXVI, pp. 2502-2507.
Hsu, T.C., Chromosome evolution in cell populations, in ‟International review of cytology", 1961, XII, pp. 69-78.
Iannaccone, P. M., Gardner, R. L., Harris, H., The cellular origin of chemically induced tumors, in ‟Journal of cell sciences", 1978, XXIX, pp. 249-255.
Illmansee, K., Mintz, B., Totipotency and normal differentiation of single teratocarcinoma cell cloned by injection into blastocysts, in ‟Proceedings of the National Academy of Sciences, USA", 1976, LXXIII, pp. 549-553.
Kerbel, R.S., Tumor heterogeneity, invsion and metastasis, in ‟Invasion metastasis", 1982, II, pp. 61-75.
Klein, E., Klein, G., A system for the detection of single gene mutations in mouse tumors, in Proceedings of the II International symposium of mammary cancer (a cura di L. Severi), Perugia 1958, pp. 709-712.
Klein, E., Klein, G., Hellstrom, K. E., Further studies on isoantigenic variation in mouse carcinomas and sarcomas, in ‟Journal of the National Cancer Institute", 1960, XXV, pp. 271-280.
Klein, G., Genetics of somatic cells, in Methodology in mammalian genetics (a cura di W. G. Burdette), San Francisco 1963, pp. 407-468.
Klein, G., Klein, E., Variation in cell population of transplanted tumors as indicated by studies on the ascites transformation, in ‟Experimental cell research", suppl., 1955, III, pp. 218-230.
Klein, G., Klein, E., Conversion of solid neoplasms into ascites tumors, in ‟Annals of the New York Academy of Sciences", 1956, LXIII, pp. 640-661.
Klein, G., Klein, E., Some experiments on the mechanism of progression in mouse mammary carcinomas, in Proceedings of the II International symposium on mammary cancer (a cura di L. Severi), Perugia 1958, pp. 713-717.
Koller, P. C., Chromosome behavior in tumors: readjustments to Boveri's theory, in Cell physiology of neoplasia, Austin, Tex., 1960, pp. 9-48.
Koller, P.C., The role of chromosomes in cancer biology, in ‟Cancer research", 1972, XXXVIII, pp. 14-56.
Lee, R.W., Gillham, N. W., Van Winkle, K. P., Boynton, J. E., Preferential recovery of uniparental streptomycin resistant mutants from diploid Chlamydomonas Reinhardti, in ‟Molecular genetics", 1973, CXXI, pp. 109-113.
Liotta, L.A., Rao, C. N., Barsky, S. N., Laminin receptors on human breast carcinoma: role in invasion of the extracellular matrix, in Understanding breast cancer: clinical and laboratory concepts (a cura di M.A. Rich, J.C. Hager e P. Furmanski), New York 1983, pp. 87-97.
Makino, S., Further evidence favoring the concept of the stem cell in ascites tumors of rats, in ‟Annals of the New York Academy of Sciences", 1956, LXIII, pp. 818-830.
Makino, S., The chromosome cytology of the ascites tumors of rats with special reference to the concept of the stemline cell, in ‟International review of cytology", 1957, VI, pp. 25-32.
Medina, D., Preneoplastic lesions in mouse mammary tumorigenesis, in ‟Methods in cancer reesarch", 1973, VII, pp. 3-15.
Medina, D., Biologic characteristics of tumor progression in the mouse mammary gland, in ‟Cancer chemotherapy reports", 1975, V, pp. 127-131.
Medina, D., Tumor progression, in Cancer (a cura di F. F. Becker), vol. III, New York 1975, pp. 99-119.
Medina, D., Regulation of expression of the preneoplastic cell phenotype, in Breast cancer research conference. Proceedings (a cura di M. A. Rich), Denver 1983, p. 22.
Medina, D., DeOme, K. B., Influence of mammary tumor virus on the tumor-producing capabilities of nodule outgrowths free of maammary tumor virus, in ‟Journal of the National Cancer Institute", 1968, XL, pp. 1303-1308.
Medina, D., DeOme, K.B., Carcinogen-induced mammary tumors from preneoplastic nodule outgrowths in BALB/c mice, in ‟Cancer research", 1970, XXX, pp. 1055-1060.
Medina, D., Shepard, F., Gropp, T., Enhancement of the tumorigenicity of preneoplastic mammary nodule lines by enzymatic dissociation, in ‟Journal of the National Cancer Institute", 1978, LX, pp. 1121-1126.
Meins, F., Reversal of the neoplastic state in plants, in ‟American journal of pathology", 1977, LXXXIX, pp. 687-702.
Miller, B. E., Miller, F. R., Leith, J., Heppner, G. H., Growth interaction in vivo between tumor subpopulations derived from a single mouse mammary tumor, in ‟Cancer research", 1980, XL, p. 3977-3981.
Miller, F.R., Miller, B.E., Heppner, G. H., Characterization of metastatic heterogeneity among subpopulations of a single mouse mammary tumor: heterogeneity in phenotypic stability, in ‟Invasion metastasis", 1983, III, pp. 22-31.
Mintz, B., Illmansee, K., Normal genetically mosaic mice produced from malignant teratocarcinoma cells, in ‟Proceedings of the National Academy of Sciences, USA", 1975, LXXII, pp. 3585-3589.
Mintz, B., Slemmer, G., Gene control of neoplasia. I. Genotypic mosaicism in normal and preneoplastic mammary glands of allophenic mice, in ‟Journal of the National Cancer Institute", 1969, XLIII, pp. 87-95.
Nandi, S., Models for transformation of breast cells in animals and man, in Abstracts of the 11th meeting on mammary cancer in experimental animals and man (a cura di M. J. Brennan e M. A. Rich), Detroit 1978, p. 25.
Nenci, I., Estrogen receptor cytochemistry in human breast cancer: status and prospects, in ‟Cancer", 1981, XLVIII, pp. 2674-2686.
Nowell, P.C., The clonal evolution of tumor cell populations. Acquired genetic lability permits stepwise selection of variant sublines and underlies tumor progression, in ‟Science", 1976, CXCIV, pp. 23-28.
Nuti, M., Castagna, M., Squartini, F., Caratterizzazione morfologica della reattività agli anticorpi monoclonali B 6.2, B 72.3 e B 1.1 nella displasia e nel cancro della mammella, in Progressi in anatomia patologica (a cura di L. Severi), Perugia 1984, pp. 91-105.
Pitot, H.C., The natural history of neoplasia: newer insights into an old problem, in ‟American journal of pathology", 1977, LXXXIX, pp. 401-412.
Pitot, H.C., The stability of the neopalstic state. Introduction, in ‟American journal of pathology", 1977, LXXXIX, pp. 668-670.
Pitot, H.C., The natural history of neoplastic development: the relation of experimental models to human cancer, in ‟Cancer", 1982, XLIX, pp. 1206-1211.
Pitot, H.C., Sirica, A.E., The stages of initiation and promotion in hepato-carcinogenesis, in ‟Biochimica et biophysica acta", 1980, DCV, pp. 191-215.
Poste, G., Tumor cell heterogeneity and the metastatic process, in Understandig breast cancer: clinical and laboratory concepts (a cura di M. A. Rich, J. C. Hager e P. Furmanski), New York 1983, pp. 119-143.
Poste, G., Doll, J., Fidler, I. J., Interactions between clonal subpopulations affect the stability of the metastatic phenotype in polyclonal populations of B 16 melanoma cells, in ‟Proceedings of the National Academy of Sciences, USA", 1981, LXXVIII, pp. 6226-6230.
Poste, G., Fidler, I. J., The pathogenesis of cancer metastasis, in ‟Nature", 1980, CCLXXXIII, pp. 139-146.
Poste, G., Greig, R., On the genesis and regulation of cellular heterogeneity in malignant tumors, in ‟Invasion metastasis", 1982, II, pp. 137-176.
Prehn, R.T., Tumor progression and homeostasis, in ‟Advances in cancer research", 1976, XXIII, pp. 203-236.
Reddy, A.L., Fialkow, P. J., Multicellular origin of fibrosarcomas in mice induced by the chemical carcinogen 3-methylcholanthrene, in ‟Journal of experimental medicine", 1980, CL, pp. 876-886.
Rubin, H., Is somatic mutation the major mechanism of malignant transformation?, in ‟Journal of the National Cancer Institute", 1980, LXIV, pp. 995-1000.
Sakakura, T., Epithelial-mesenchimal interactions in mammary gland development and its perturbation in relation to tumorigenesis, in Understanding braest cancer: clinical and laboratory concepts (a cura di M. A. Rich, J.C. Hager e P. Furmanski), New York 1983, pp. 261-284.
Silagi, S., Modification of malignancy by 5-bromodeoxyuridine. Studies of reversibility and immunological effects, in ‟In vitro", 1971, VII, pp. 105-114.
Silagi, S., Reversible suppression of malignancy and differentiation of melanoma cells, in ‟American journal of pathology", 1977, LXXXIX, pp. 671-684.
Sinha, D., Dao, T. L., A direct mechanism of mammary carcinogenesis induced by 7,12-dimethylbenzanthracene, in ‟Journal of the National Cancer Institute", 1974, LIII, pp. 841-846.
Slemmer, G. L., Interaction of separate types of cells during normal and neoplastic mammary gland growth, in ‟Journal of investigative dermatology", 1974, LXIII, pp. 27-47.
Slemmer, G.L., Interactions of different types of mammary cells during progression and metastasis of breast neoplasia, in Cell biology of breast cancer (a cura di C. M. McGrath, M. A. Rich e M. J. Brennan), New York 1980, pp. 93-143.
Sporn, M. B., Approaches to prevention of epithelial cancer during the preneoplastic period, in ‟Cancer rsearch", 1976, XXXVI, pp. 2699-2702.
Squartini, F., III. Histogenesis, in Brest cancer research. UICC technical report series (a cura di M. J. Brennan), vol. XXVII, Genève 1977, pp. 51-58.
Squartini, F., Tumours of the mammary gland, in Pathology of tumours in laboratory animals, vol. II, Tumours of the mouse (a cura di V.S. Turusov), Lyon 1979, pp. 43-90.
Squartini, F., Lesioni preneoplastiche sperimentali e umane della mammella, in Apporto della ricerca di base al controllo della crescita neoplastica (a cura di F. Bresciani), Napoli 1981, pp. 397-404.
Squartini, F., MuMTV (murine mammary tumor virus) e carcinoma mammario, in Virus oncogeni ad RNA (Retrovirus) (a cura di C. De Giuli Morghen), Padova 1981, pp. 228-276.
Squartini, F., Lesioni preneoplastiche e fasi precoci del cancro mammario sperimentale e umano, in ‟Aggiornamento del medico", 1982, I, pp. 49-62.
Squartini, F., Storia naturale del cancro della mammella: la progressione tumorale, in ‟Acta oncologica", 1982, III, pp. 119-124.
Squartini, F., The mammary lobule in cancerous breast: human findings and experimental background, in New frontiers in mammary pathology (a cura di K. H. Hollman e J. M. Verley), vol. II, New York 1983, pp. 165-209.
Squartini, F., Basolo, F., Bistocchi, M., Lobuloalveolar differentiation and tumorigenesis: two separate activities of mouse mammary tumor virus, in ‟Cancer research", 1983, XLIII, pp. 5879-5882.
Squartini, F., Bistocchi, M., Bioactivity of C3H and RIII mammary tumor viruses in virgin female BALB/c mice, in ‟Journal of the National Cancer Institute", 1977, LVIII, pp. 1845-1847.
Squartini, F., Bistocchi, M., Buongiorno, L., Development, morphology and progression of mammary tumors during and after fertile life in BALB/cfRIII mice, in ‟Journal of the National Cancer Institute", 1981, LXVI, pp. 311-319.
Squartini, F., Pingitore, R., Bistocchi, M., Characterization of RIII mouse mammary tumor virus, in Breast cancer. New concepts in etiology and control (a cura di M. J. Brennan, C. M. McGrath e M. A. Rich), New York 1980, pp. 213-223.
Squartini, F., Severi, L., Strain differences in the mammary tumour-inducing virus as detected by the characters and behaviour of neoplasms, in A Ciba Foundation symposium on tumor viruses of murine origin (a cura di G.E.W. Wolstenholme e M. O'Connor), London 1962, pp. 82-106.
Stock, J. A., Chemioterapia dei tumori, in Biologia dei tumori (a cura di J. Ambrose e F. J. C. Roe), Napoli 1978, pp. 315-354.
Sugarbaker, E.V., Some characteristics of metastasis in man, in ‟American journal of pathology", 1979, XCVII, pp. 623-632.
Talmadge, J. E., The selective nature of metastasis, in ‟Cancer metastasis reviews", 1983, II, pp. 25-40.
Talmadge, J. E., Starkey, J. R., Stanford, D.R., In vitro characteristics of metastatic variant subclones of restricted genetic origin, in ‟Journal of supramolecular and structural cell biochemistry", 1981, XV, pp. 139-151.
Tanouka, H., Tanaka, K., Evidence for single cell origin of 3-methylcholanthrene-induced fibrosarcoma in mice with cellular mosaicism, in ‟Cancer research", 1982, XLII, pp. 1857-1858.
Weinberg, R.A., Oncogeneses of spontaneous and chemically induced tumors, in ‟Advances in cancer research", 1982, XXXVI, pp. 149-163.
Weiss, L., Dynamic aspects of cancer cell populations in metastasis, in ‟American journal of pathology", 1979, XCVII, pp. 601-608.
Willis, R.A., Pathology of tumours, London 19674.
Woodruff, M. F. A., Ansell, J.D., Forbes, G. M., Gordon, J. C., Burton, D. I., Micklem, H.S., Clonal interaction in tumors, in ‟Nature", 1982, CCXCIX, pp. 822-824.
Dolore e terapia del dolore
di Umberto Veronesi, Franco De Conno, Michele Gallucci, Vittorio Ventafridda
sommario: 1. Introduzione: a) definizione e generalità; b) significato del dolore; c) il dolore come malattia. 2. Elementi di anatomia e di fisiologia: a) recettori del dolore; b) vie di conduzione e centri della sensibilità dolorifica; c) integrazioni della sensibilità dolorifica; d) la percezione del dolore nell'uomo; e) il dolore centrale. 3. Il dolore in clinica oncologica: a) importanza del problema; b) incidenza del dolore nelle neoplasie; c) cause del dolore nelle neoplasie; d) problemi psicologici del dolore; e) semeiologia e diagnosi del dolore. 4. Terapia del dolore: a) strategia terapeutica; b) terapia oncologica specifica; c) terapia di lesione; d) terapia di modulazione; e) unità di assistenza continua. 5. Conclusioni. □ Bibliografia.
1. Introduzione
a) Definizione e generalità
La definizione del dolore, a prescindere dalle cognizioni acquisite con gli studi elettrofisiologici e psicofisici, nasce dalla comune esperienza di ogni essere cosciente: si tratta di una sensazione nettamente e più o meno fortemente sgradevole, localizzabile in una zona della superficie cutanea, o in profondità, o nei visceri. Peraltro, esaminando più attentamente il complesso di tale sensazione, è possibile individuarne due componenti: una sensitiva, rappresentata dalla percezione immediata dello stimolo algogeno e della sua intensità e poco o affatto influenzata dall'emotività; e una psichica, rappresentata dalle reazioni di ordine affettivo, per solito assai bene evidenti nei dolori che accompagnano i processi morbosi, variabile nei vari individui. È questo secondo aspetto dell'esperienza sensitiva dolorosa che, sommandosi alle reazioni vasomotorie e neurovegetative che ne conseguono e che entrano a loro volta a farne parte, dà luogo a quella particolare condizione emotiva genericamente indicata come sofferenza.
La capacità di percepire gli stimoli dolorosi rappresenta per l'organismo un valido segnale dell'esistenza di un pericolo per la propria integrità o di una condizione patologica in evoluzione. Tuttavia, mentre nel primo caso, così frequente ad esempio per gli stimoli meccanici e termici di una certa intensità agenti sulla cute, la reazione scatenata dalla sensazione dolorosa si traduce nell'allontanamento dall'agente lesivo, il dolore provocato da un viscere malato persiste invece per tutta la durata del processo morboso. Si comprende facilmente, quindi, quali problemi possano prospettare malattie croniche, a lungo decorso, nelle quali il dolore può talvolta assorbire tutta l'attenzione del paziente e divenire l'elemento dominante nell'evoluzione del processo morboso.
b) Significato del dolore
Le considerazioni sopra esposte sulle origini e sulle componenti del dolore consentono di individuarne vari significati dal punto di vista biologico e da quello clinico (v. Porta, 19762). S'è detto che uno stimolo algogeno che viene percepito a livello di una determinata zona cutanea provoca l'allontanamento o la fuga: se questa reazione, valida a preservare l'integrità dell'organismo, avviene ancora prima che l'agente nocivo eserciti una lesione a carico delle cellule, il dolore costituisce un semplice segnale, una pura sensazione le cui componenti percettiva ed emotiva sono limitate nel tempo.
Se l'agente nocivo che è all'origine dello stimolo algogeno provoca un danno cellulare, si determinano liberazione locale di sostanze vasoattive e successive alterazioni flogistiche che costituiscono un ulteriore stimolo doloroso: la sensazione che ne consegue è allora più complessa, persiste più a lungo, può accompagnarsi a reazioni generali neurovegetative e psichiche. Il dolore, in tal caso, acquista il significato di sintomo della lesione provocata dall'agente nocivo, pur conservando il valore biologico di segnale capace di provocare l'allontanamento o la fuga e di limitare in tal modo il danno subito dall'organismo.
Quando lo stimolo algogeno deriva dalla profondità dell'organismo, ad es. da un viscere malato, il dolore conserva esclusivamente il significato di sintomo, in quanto, essendo spontaneo, in nessun caso può dare origine a reazioni capaci di allontanare l'agente nocivo. È bensì vero che il valore generico di allarme in grado di richiamare l'attenzione sull'organo malato, che gli si attribuisce, può inizialmente essere utilizzato ai fini diagnostici, ma è indubbio che in tali condizioni il dolore acquista il significato di gran lunga dominante di sintomo, talvolta penoso e intollerabilmente persistente.
c) Il dolore come malattia
Si designa propriamente col termine di ‛dolore malattia' il dolore correlato a condizioni irritative dei centri nervosi deputati all'elaborazione degli impulsi afferenti (v. Porta, 19762). Tuttavia, in senso più generale sembra possibile estendere il concetto e il termine di dolore malattia ad altre situazioni patologiche, in particolare a molti casi di neoplasie, in cui il dolore esclusivamente sintomo, cui si è fatto cenno, raggiunge livelli così elevati di intensità da assorbire interamente l'attenzione del malato e determinarne uno stato di coscienza assolutamente peculiare. È proprio in questi casi che la penosissima percezione della sensazione dolorosa primaria e le reazioni che a questa fanno seguito danno origine a quell'esperienza sensitiva così complessa in cui una gran parte è rappresentata dalla tonalità emotiva. In questi casi il dolore, che meglio potrebbe forse essere definito sofferenza angosciosa, trascende qualunque significato biologico e fisiologico e diviene l'elemento dominante del quadro clinico, che solo esige, in qualunque modo, il sollecito intervento terapeutico; il dolore, in altri termini, può essere considerato come una vera e propria malattia.
La ‛malattia dolore' del paziente portatore di una neoplasia, che giustifica l'impiego di determinate strategie terapeutiche volte spesso solo ed esclusivamente a eliminare la sofferenza senza minimanente agire sulla causa che la determina, costituisce il campo di applicazione del nuovo, grande capitolo della clinica oncologica: la terapia del dolore.
2. Elementi di anatomia e di fisiologia
a) Recettori del dolore
Si designano genericamente col termine di recettori le terminazioni periferiche delle fibre nervose afferenti con le loro formazioni accessorie; queste strutture, in quanto dotate di specifica sensibilità, sono capaci di reagire a stimoli adeguati con fenomeni di depolarizzazione da cui originano gli impulsi nervosi propagati lungo le fibre sensitive (v. elettrofisiologia; v. neurone e impulso nervoso; v. recettori; v. somatoestesia). Le proprietà peculiari dei recettori sono quindi l'eccitabilità, cioè la capacità di dare origine a un potenziale d'azione la cui ampiezza è in relazione esponenziale con l'intensità dello stimolo, e l'adattamento, cioè la capacità di mantenere e stabilizzare la scarica che dà origine all'impulso nervoso. L'eccitabilità delle terminazioni nervose sensitive è caratterizzata da un determinato valore di soglia, per cui gli stimoli adeguati risultano efficaci soltanto quando raggiungono una certa intensità. L'adattamento, individuabile nel declino del potenziale generatore durante una stimolazione prolungata, si sviluppa con differente velocità nei vari tipi di recettori: rapidamente in quelli detti appunto rapidi o fasici, che cessano di rispondere dopo un breve periodo di stimolazione; lentamente in quelli detti lenti o tonici, la cui risposta si prolunga con la persistenza dello stimolo.
I recettori del dolore, o nocicettori, sono recettori ubiquitari a lento adattamento, a soglia elevata, connessi a piccoli campi recettivi, in grado di reagire a stimoli tattili (meccanonocicettori) e termici (termonocicettori) di una certa intensità. È stato dimostrato che i nocicettori sono sensibili a numerose sostanze chimiche (istamina, serotonina, acetilcolina, ioni potassio, adenosintrifosfato, prostaglandine, bradichinina, sostanza P), molte delle quali si liberano dalle cellule o passano nel tessuto interstiziale dai vasi sanguigni in conseguenza del danno causato dall'agente nocivo; di tali sostanze alcune sono in grado di agire direttamente sulle terminazioni nervose, altre agiscono come intermediari. Sembra che la sostanza P, un undecapeptide prodotto da cellule del sistema nervoso centrale e periferico, oltre ad agire come sostanza algogena periferica, svolga il ruolo di neuromodulatore a livello delle afferenze primarie di senso (v. Barker, 1977; v. neurosecrezione; v. somatoestesia). I nocicettori sono quindi sensibili a tutti gli agenti nocivi esterni, che pertanto producono stimoli algogeni superficiali, e a fenomeni che si svolgono nell'interno dell'organismo, quali flogosi e ischemia dei tessuti, distensione di visceri cavi, vasocostrizione, ecc.
b) Vie di conduzione e centri della sensibilità dolorifica
Gli impulsi algogeni generati nei nocicettori vengono propagati lungo i nervi periferici attraverso le fibre del gruppo A delta, mielinizzate e a conduzione rapida, e del gruppo C, amieliniche e a conduzione lenta. Gli studi elettrofisiologici hanno consentito di dimostrare che, a causa della differente velocità di conduzione dei due tipi di fibre, ogni stimolo dolorifico periferico produce una sensazione immediata, vivida e circoscritta (first pain o fast pain) seguita a breve distanza di tempo da una seconda sensazione, meno precisa e localizzata, più diffusa e sgradevole (second pain o slow pain). Le fibre che conducono gli stimoli dolorifici penetrano nel ganglio spinale; di qui la branca ascendente del protoneurone, attraverso la radice dorsale, giunge nel midollo, vi entra, risale per alcuni mielomeri e penetra nelle corna grigie posteriori, ove contrae sinapsi con piccole cellule della sostanza gelatinosa di Rolando e con i grossi sensoneuroni del quarto, quinto e sesto strato. Su ogni sensoneurone convergono più fibre afferenti somatiche e viscerali. I neuriti di tali neuroni si incrociano sulla linea mediana, raggiungono il fascio spinotalamico laterale del lato opposto e pervengono al nucleo lateroventrale posteriore del talamo, costituendo pertanto una via sensitiva oligosinaptica. Un contingente dei neuriti del secondo neurone dà invece origine a una via sensitiva multisinaptica, prendendo connessione, prima di raggiungere il talamo, con neuroni della sostanza reticolare da cui hanno origine vie sensitive di terzo ordine.
Dal talamo si dipartono fibre che, attraverso il braccio posteriore della capsula interna, si distribuiscono diffusamente a tutta la corteccia cerebrale, probabilmente senza una precisa corrispondenza somatotopica. Il complesso giuoco di connessioni che si stabiliscono a vario livello tra vie di proiezione della sensibilità dolorifica e centri superiori è responsabile da un lato della formazione dell'esperienza sensitiva dolorosa con le sue componenti percettiva e affettiva, dall'altro dello sviluppo delle reazioni vasomotorie e neurovegetative che entrano a farne parte e dei fenomeni comportamentali che ne conseguono.
c) Integrazioni della sensibilità dolorifica
A livello midollare e cerebrale si stabiliscono importanti connessioni responsabili del meccanismo di integrazione del dolore (v. Porta, 19762). Studi citoarchitettonici (v. Rexed, 1954) e ultrastrutturali (v. Ralston, 1968) condotti nel gatto hanno consentito di dimostrare che nel corno posteriore del midollo spinale sono riconoscibili sei strati cellulari. Lo strato 1 contiene neuroni che rispondono solo a stimoli molto intensi e vengono inibiti dalla stimolazione delle aree cutanee confinanti con le corrispondenti terminazioni periferiche. Gli strati 2 e 3 corrispondono alla cosiddetta sostanza gelatinosa. Nello strato 5 sono contenute grosse cellule che rispondono a stimolazioni a soglia elevata e si proiettano su neuroni situati ventralmente, i cui assoni vanno ai cordoni dorsolaterali omo- e controlaterali. Su queste cellule esercitano facilitazione dello stato eccitatorio gli impulsi nocicettivi e inibizione quelli a bassa soglia provenienti sia dal tessuto stimolato (che in una breve fase iniziale sono invece eccitatori), sia dalle zone a questo circostanti; inoltre esercitano azione inibente della risposta agli stimoli afferenti algogeni le stimolazioni piramidale, rubrospinale e reticolospinale. La capacità di queste cellule di rispondere ad afferenze viscerali e cutanee dello stesso dermatomero spiega l'origine del cosiddetto dolore riferito o eterotopico (v. § d). Queste acquisizioni costituiscono la base anatomica della teoria del cosiddetto gate control system: sulla cellula bersaglio T, identificabile in un grosso sensoneurone dello strato 5, agiscono impulsi afferenti da fibre radicolari A, a bassa soglia e a rapido adattamento, e da fibre radicolari C, a soglia elevata e a lento adattamento. Il neurone T è dapprima eccitato dagli impulsi provenienti da A, poi inibito mediante un circuito ricorrente negativo, forse implicante le piccole cellule della sostanza gelatinosa che sono inibitorie presinaptiche; gli impulsi provenienti da C esercitano su T un effetto facilitante per inibizione delle cellule della sostanza gelatinosa, che come s'è detto sono inibitorie, cosicché l'azione stimolante degli impulsi afferenti ne risulta esaltata. Gli impulsi afferenti dalle fibre A tendono a ‛chiudere lo sportello', quelli provenienti dalle fibre C ad ‛aprirlo' rispettivamente deprimendo o aumentando l'attività del sensoneurone T. Lo ‛sportello' è anche controllato sia da impulsi che discendono sul neurone T dal cervelletto, dal talamo, dal nucleo rosso, dalla corteccia cerebrale, sia dal sistema reticolare, per l'esistenza di connessioni polisinaptiche spinoreticolari e reticolospinali. Gli impulsi discendenti sul sensoneurone dai centri superiori sono a loro volta attivati da sistemi afferenti a conduzione rapida dei cordoni posteriori. Fanno parte dell'integrazione midollare del dolore anche le reazioni riflesse, indipendenti dai centri superiori, tendenti ad allontanare lo stimolo nocicettivo.
L'esistenza di un sistema di controllo delle afferenze nocicettive localizzato in determinate sedi encefaliche, soprattutto tronco dell'encefalo e talamo, è stata ipotizzata a seguito di due fondamentali osservazioni: la possibilità di indurre analgesia mediante stimolazione elettrica delle fibre afferenti e la dimostrazione di molecole recettrici per gli oppiacei nella membrana di gruppi di neuroni di regioni specifiche dell'encefalo (v. somatoestesia). Le regioni che, stimolate elettricamente, producono riduzione o scomparsa del dolore sono situate sia nel sistema somatico afferente - come le terminazioni nervose cutanee, le fibre periferiche, le colonne dorsali e ventrali, il complesso talamico ventrobasale, la capsula interna - sia nella sostanza grigia periventricolare e periacqueduttale del talamo e del mesencefalo nonché nel nucleus magnus del rafe mediano (v. Long e Frickson, 1975; v. Fields e Basbaum, 1978). Recettori neuronali per gli oppiacei sono stati localizzati a livelli in gran parte corrispondenti alle zone la cui stimolazione elettrica è in grado di indurre analgesia: si è visto infatti che piccole quantità di morfina iniettate nel corno posteriore del midollo spinale o in certe regioni dell'encefalo, specie nella sostanza grigia periacqueduttale e periventricolare, producono analgesia (v. Yaksh e Rudy, 1978). Inoltre nell'encefalo sono stati scoperti neuropeptidi capaci di interagire con i recettori membranari per gli oppiacei; questi peptidi, chiamati encefaline ed endorfine e derivanti tutti dalla beta-lipotropina, una proteina presente nell'ipofisi anteriore e intermedia e in cellule dell'ipotalamo (v. Hughes, 1975; v. Hughes e altri, 1975; v. Klee, 1977; v. Miller, 1978), potrebbero esercitare un controllo naturale del dolore comportandosi come neuromodulatori.
A livello cerebrale si stabiliscono le importanti connessioni tra vie e centri della sensibilità dolorifica e meccanismi regolatori neurovegetativi. Le proiezioni nelle aree sensitive corticali offrono la rappresentazione dello schema corporeo che consente di localizzare il dolore, mentre le vie talamofrontali sono responsabili del controllo della sensazione dolorosa come esperienza del tutto personale. La corteccia quindi partecipa con fenomeni di facilitazione e di inibizione alla percezione del dolore non limitatamente all'area sensitiva; la funzione dei complessi sistemi polineuronici che costituiscono la via centrale del dolore sembra pertanto quella di integrare gli impulsi dolorifici con le condizioni globali dell'organismo (v. Porta, 19762). Pur non essendo possibile stabilire con certezza dove si formi la sensazione dolorosa, sembra evidente che nel talamo si produca la sensibilità protopatica, cioè la coscienza affettiva dell'esperienza dolorosa, mentre a livello corticale avrebbe luogo la percezione epicritica del dolore, cioè la sua localizzazione in una determinata zona corporea e la valutazione del tipo e dell'intensità dello stimolo algogeno stesso.
d) La percezione del dolore nell'uomo
L'uomo è in grado di percepire una gamma di sensazioni dolorose essenzialmente riconducibili a tre tipi: dolore superficiale, dolore profondo, dolore viscerale.
Il dolore superficiale è quello prodotto, ad esempio, da uno stimolo nocivo che agisce sulla cute: si tratta di una sensazione ben distinta e localizzata, che cessa quando viene rimosso lo stimolo. È esperienza comune che uno stimolo algogeno di una certa intensità agendo sulla cute dà origine a due distinte sensazioni dolorose: una prima acuta e di breve durata, seguita, dopo un piccolo intervallo di tempo, da una seconda più intensa, più lunga, meno precisa. Queste due sensazioni corrispondono alle risposte dei due differenti tipi di terminazioni nervose, A delta e C, a diversa velocità di conduzione, cui s'è fatto cenno in precedenza.
Il dolore profondo è non precisamente localizzato, non ben distinto, sordo; insorge dopo una certa latenza e può persistere a lungo, anche quando lo stimolo cessa di agire. Esso si manifesta per alterazioni delle strutture profonde - muscoli, fasce, tendini, articolazioni, periostio ed è solitamente caratterizzato dall'apparente spontaneità.
Il dolore viscerale è prodotto dalla distensione dei visceri cavi, dalla costrizione dei vasi sanguigni, dalla trazione e distorsione delle sierose parietali; gli organi parenchimatosi, come fegato, polmoni, rene, milza, non sono sensibili alle varie stimolazioni. Il dolore viscerale che origina dalla vescica e dagli organi pelvici viene propagato lungo le fibre del parasimpatico coccigeo, quello che origina dai visceri addominali e toracici viene propagato lungo le fibre simpatiche dei nervi e dei plessi periarteriosi. La sofferenza di origine viscerale può essere distinta in un dolore viscerale vero o splancnico, percepito nel viscere stesso, piuttosto diffuso e poco localizzato, e in un dolore parietale, più superficiale, acuto e precisamente localizzato, dovuto alla flogosi reattiva aspecifica delle sierose interessate nel processo morboso, che si accompagna a contrattura della parete. Il dolore parietale può essere percepito a distanza dal punto di origine in conseguenza di contratture riflesse che si stabiliscono quando la stimolazione algogena agisca su strutture provviste di varie innervazioni: così, ad esempio, una flogosi pericardica può indurre per contiguità sofferenza del diaframma, il quale può di riflesso contrarsi e anche scatenare contrazioni riflesse dei muscoli intercostali, in quanto l'innervazione periferica del diaframma è assicurata dai nervi intercostali.
Il dolore viscerale può essere percepito in una zona cutanea distante dall'organo da cui origina: è questo il cosiddetto dolore proiettato, o a distanza, o eterotopico, o riferito, che può accompagnarsi a quello originato dal viscere ove agisce lo stimolo nocivo o manifestarsi in completa assenza di questo (v. anche somatoestesia). Il dolore eterotopico interessa la zona centrale dei dermatomeri corrispondenti ai mielomeri che ricevono afferenze dal viscere in cui agisce lo stimolo nocivo; uno dei più noti dolori eterotopici è quello irradiato al lato ulnare dell'arto superiore sinistro in corso di crisi stenocardica. Il dolore eterotopico si spiega sia tenendo presente che su uno stesso sensoneurone si articolano fibre viscerali e fibre somatiche, così che afferenze algogene viscerali possono ripercuotersi somaticamente alla periferia rispettando la distribuzione dermatomerica; sia con la possibilità che le stimolazioni algogene viscerali giungano attraverso fibre collaterali ai sensoneuroni somatici, la cui conseguente subeccitazione sia in grado di determinare un abbassamento della soglia per le stimolazioni cutanee. A dimostrazione di ciò sta la constatazione che in caso di stimolazione algogena viscerale di lieve entità si può ottenere la scomparsa del dolore eterotopico anestetizzando la cute della zona di riferimento o le fibre che la innervano.
e) Il dolore centrale
Molte condizioni morbose che interessano il sistema nervoso - dalle lesioni delle corna posteriori alla siringomielia, alla sclerosi multipla, ad alcune neoplasie - determinano dolori di elevata intensità che insorgono spontaneamente o a seguito di stimoli abitualmente innocui. Espressione paradigmatica di questo tipo di dolore è quello peculiare della sindrome talamica di Déjerine e Roussy (v., 1906): si tratta di un'algia intollerabile, diffusa alla metà del corpo controlaterale alla lesione, continua o subcontinua, esacerbata dalle più lievi stimolazioni tattili o termiche. La soglia del dolore può essere addirittura elevata, ma non appena la raggiunge lo stimolo scatena la sofferenza. Si tratta del dolore malattia cui si è precedentemente accennato (v. cap. 1, § c). L'origine del dolore centrale va ricercata nelle alterazioni sia delle interazioni talamocorticali con dissociazione della sensibilità protopatica da quella epicritica, sia della funzione reticolare normalmente esercitantesi sull'attività cerebrale cosciente e sugli stati emotivi.
3. Il dolore in clinica oncologica
a) Importanza del problema
Pur se da molto tempo è noto che il dolore cronico colpisce la maggior parte dei pazienti portatori di neoplasie, costituendo, soprattutto nelle fasi terminali, quell'evento fondamentale in grado di alterare la qualità di vita che si è genericamente definito ‛malattia dolore' (v. cap. 1, § c), gli sforzi della clinica oncologica per ricercare la soluzione di tale problema hanno un inizio relativamente recente. La limitata informazione, soprattutto per quanto riguarda la raccolta e l'elaborazione statistica dei dati, e un certo vuoto scientifico intorno all'argomento possono originare da varie cause, che tenteremo ora di analizzare.
Anzitutto occorre rilevare che il numero di pubblicazioni specializzate è scarso: in undici testi fondamentali sulle neoplasie, soltanto 21 pagine su 9.000 sono dedicate al trattamento del dolore (v. Bonica e Ventafridda, 1979); in particolare nella letteratura medica di lingua inglese il problema è scarsamente trattato o del tutto ignorato. Praticamente negli ultimi trent'anni non è stata pubblicata alcuna ricerca sui meccanismi del dolore da neoplasia, a conferma che l'interesse degli studiosi è concentrato esclusivamente sulla fisiopatologia generale del dolore.
Vi è poi un'allarmante carenza di fondi destinati a tale ricerca: negli Stati Uniti viene annualmente stanziata per studi sul dolore una somma pari allo 0,039% del miliardo di dollari destinato alla ricerca di nuove armi contro i tumori; nel prestigioso Nati onal Institute of Health viene destinato alle ricerche sul dolore da neoplasie solo lo 0,02% dei fondi.
L'interesse mostrato dagli studiosi per il problema è generalmente scarso: in tutti gli Stati Uniti non più di dieci scienziati si dedicano allo studio dei meccanismi del dolore nelle neoplasie, ed è esperienza comune che i protocolli terapeutici, i risultati clinici, i programmi di informazione didattica nella lotta contro i tumori ignorano ed eludono la parola dolore.
L'educazione sanitaria nei riguardi del problema è assai carente o manca del tutto: a livello universitario o specialistico mancano i corsi di informazione e gli accenni alle modalità e ai meccanismi del dolore da neoplasie nonché alle risposte fisiologiche emozionali e comportamentali al dolore sono rudimentali.
Infine piuttosto scarse sono le conoscenze farmacologiche sul dolore: la farmacologia clinica del dolore cronico non ha realizzato grandi progressi negli ultimi anni, ove si eccettuino gli studi condotti nell'ultimo decennio sui neuropeptidi endogeni (v. cap. 2, § c).
b) Incidenza del dolore nelle neoplasie
I dati desunti a partire dalle prime osservazioni, risalenti al 1954, fino alle revisioni più recenti (v. Twycross e Ventafridda, 1980) mettono in evidenza che la frequenza del dolore nelle neoplasie in fase avanzata riportata nelle casistiche da vari autori va dal 60 all'80% circa (v. tab. I). Questi dati, pur se non omogenei e non sottoposti a esame critico, configurano una realtà clinica drammatica.
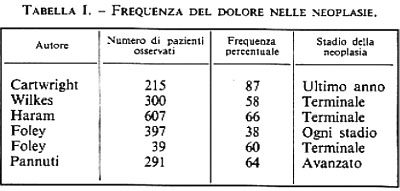

Assumendo come valide tali percentuali e proiettandole nella clinica oncologica italiana, è possibile stimare che ogni anno in Italia su 250.000 pazienti portatori di neoplasie maligne ne muoiano 110.000, e di questi da 55.000 a 80.000 con dolore. Da stime simili risulta che in Gran Bretagna la metà almeno di tutti i pazienti che muoiono per neoplasia soffre di dolore e circa un ottavo di dolore grave. I dati statistici consentono inoltre di rilevare che i tumori che con maggiore frequenza causano dolore sono quelli a primitiva localizzazione ossea, mentre le leucemie sono causa di dolore solo in una scarsa percentuale di casi (v. tab. II).
c) Cause del dolore nelle neoplasie
Già si è parlato in generale del dolore e si è fatto cenno alla distinzione tra nocicezione, cioè segnalazione veicolata da fibre specializzate dell'azione di un'energia meccanica o termica potenzialmente lesiva di un tessuto, e dolore sintomo, cioè esperienza spiacevole e privata, primariamente connessa a un danno di un settore dell'organismo e descritta in termini di tale danno, la cui presenza è segnalata da qualche forma di comportamento visibile e udibile. Il dolore percepito dai portatori di neoplasia in fase avanzata è abitualmente cronico, di varia intensità, sofferto come un'esperienza che coinvolge la sfera emotiva, istintiva e affettiva, molto spesso così intensamente e totalmente da occupare tutta l'attenzione del paziente e isolano dal mondo estemo, assumendo così il ruolo di una vera malattia, la ‛malattia dolore' (v. cap. 1, § c). Il peculiare stato d'animo di un paziente in tali condizioni è determinato soprattutto dalla componente psichica della sensazione dolorosa; la consapevolezza dell'impossibilità di prevedere una fine prossima del dolore, della sua tendenza a peggiorare, dell'apparente assenza di un motivo che lo giustifichi alimenta la complessità della componente psichica stessa.
Si è potuto valutare che il dolore, nei pazienti portatori di neoplasie, nel 77% dei casi è in rapporto all'azione invasiva o compressiva tumorale, nel 19% dei casi consegue a terapie oncologiche specifiche, nel 3% dei casi è apparentemente indipendente dal tumore e dalla terapia. Una precisa elencazione delle cause fisiche del dolore nel corso di neoplasie è stata compilata da Bonica (v. tab. III; v. Bonica e Ventafridda, 1979): si tratta di cause molteplici, spesso associate, che comprendono compressioni di tronchi nervosi da parte del tumore primitivo o delle metastasi, fratture patologiche, flogosi tessutali, occlusioni di visceri cavi, costrizioni vascolari, neuropatie paraneoplastiche. Anche il dolore non direttamente causato dal tumore, ma a questo associato, come nel caso appunto della polineuropatia paraneoplastica, può presentare gli stessi caratteri di cronicità e di gravità.

Le varie condizioni elencate causano dolore con meccanismi diversi. La compressione di un tronco nervoso biocca la conduzione degli impulsi afferenti nelle fibre di calibro maggiore e dà quindi luogo a un'anestesia che a sua volta è causa di uno squilibrio delle afferenze ai sensoneuroni delle corna grigie posteriori e, in definitiva, dell'apertura dello ‛sportello del dolore' (v. cap. 2, § c): ne conseguono disturbi sensitivi e dolori da deafferentazione, parestesie, disestesie, iperalgesie, dolori parossistici.
Le metastasi ossee e le fratture patologiche da queste provocate sono causa di dolore soprattutto per la distensione e l'irritazione del periostio e per la liberazione locale di prostaglandine.
I tessuti infiltrati sono sede di flogosi e di alterazioni regressive varie fino alla necrosi: si rendono libere, in tali condizioni, quelle sostanze chimiche in grado di agire come mediatori o neuromodulatori periferici del dolore (v. cap. 2, § c), che rendono i nocicettori eccitabili anche per stimoli abitualmente non algogeni.
Nei visceri cavi l'infiltrazione neoplastica provoca liberazione di metaboliti algogeni, principalmente prostaglandine che, determinando lo spasmo delle fibre muscolari lisce, contribuiscono alla genesi del dolore.
L'ostruzione dei vasi arteriosi provoca ipossia e acidosi tessutale, quella delle vene edema e distensione dei tessuti: in ambedue le condizioni si determina liberazione dei mediatori chimici dei nocicettori. L'insieme di tali fenomeni è causa di dolore.
La polineuropatia paraneoplastica è una sindrome dolorosa periferica in rapporto a demielinizzazione delle fibre nervose con distruzione dei cilindrassi, verosimilmente conseguenti all'azione tossica esercitata da metaboliti prodotti e immessi in circolo dal tumore. Relativamente frequente è la polineuropatia periferica associata a carcinoma del polmone, che può precedere la comparsa dei segni diretti della neoplasia.
d) Problemi psicologici del dolore
I vari significati biologici, fisiopatologici e clinici del dolore, che sono stati esaminati precedentemente, hanno tutti in comune un elemento fondamentale: la consapevolezza dello stato di malessere, delle condizioni anormali in cui si trova un determinato distretto dell'organismo. Numerosi fattori concorrono nello sviluppo e nella manifestazione di tale stato di coscienza.
Anzitutto occorre considerare la soglia del dolore, che può variare nei vari soggetti ma tende a essere abbastanza stabile nello stesso soggetto. È evidente che non è possibile apprezzare una soglia nel caso di dolori viscerali o profondi, a differenza di quanto accade nel caso di stimoli nocicettivi portati, ad esempio, sulla superficie cutanea, ma è pensabile che anche le sofferenze che originano nell'interno dell'organismo siano apprezzabili nei vari individui in conformità alla reattività delle loro terminazioni nocicettive. Per se stessa la soglia del dolore dipende quindi esclusivamente dall'intensità dello stimolo algogeno e dall'irritabilità dei nocicettori. Tuttavia un abbassamento o un innalzamento della soglia del dolore possono apparentemente verificarsi in dipendenza da altri fattori, di natura prevalentemente psicologica.
Un apparente abbassamento della soglia del dolore può dipendere da una diminuita tolleranza degli stimoli algogeni: è quanto si verifica quando, per varie cause, si stabilisce un'aumentata predisposizione alla sofferenza dei centri nervosi sensitivi. Tale situazione è caratteristica di quegli stati che abbiamo genericamente definito di ‛malattia dolore', in cui la sofferenza si espande progressivamente invadendo tutta la coscienza dell'individuo, fino a focalizzarne completamente l'attenzione: questo stato di malattia dolore da diminuita tolleranza della sofferenza, tale da rappresentare l'elemento fondamentale del quadro clinico, si osserva con varia frequenza nelle neoplasie. Una diminuita tolleranza del dolore determina talvolta un'esaltazione tale della percezione dolorifica da far si che stimoli algogeni di lieve entità diventino insopportabili. Al contrario, sono ben noti casi di abnorme tolleranza del dolore, di una sorta di deviazione degli impulsi dolorifici, che non vengono più elaborati dai centri superiori e non sono quindi in grado di alterare lo stato di coscienza. La tolleranza del dolore è il risultato dell'elaborazione subita dagli impulsi nocicettivi afferenti a livello dei centri superiori in quel complesso fenomeno di correlazioni che caratterizzano l'elaborazione centrale del dolore (v. cap. 2, § c).
La tolleranza del dolore può diminuire in conseguenza di cause contingenti, di natura organica e affettiva, e quindi della stessa presenza della neoplasia. Su di essa, tuttavia, esercitano innegabili influenze elementi peculiari della personalità del paziente - socioculturali, etnici, famigliari - che sono alla base della modalità con cui questi manifesta la propria sofferenza: 1) l'espressività, cioè il modo di rispondere a uno stimolo algogeno; 2) la lamentela, cioè il modo di presentare la percezione del dolore; 3) il linguaggio, cioè il modo di comunicare e descrivere il dolore. La tolleranza del dolore è inoltre profondamente alterata da fattori di ordine puramente psicologico: 1) l'aspettativa emotiva del dolore in grado di modificarne la percezione, legata alla memoria di precedenti esperienze dolorose e alla situazione clinica attuale; 2) il livello di intelligenza e l'attenzione prestata alla sofferenza, da cui dipendono il significato che il paziente attribuisce al dolore e la possibilità di affievolirne la percezione; 3) il grado di emotività del paziente, cui sono correlate le reazioni viscerali e mimico-espressive al dolore. In questo complesso di sensazioni e di reazioni, che coinvolgono le sfere organica e psichica del paziente, è implicato, oltre al talamo e ai centri corticali superiori, anche il rinencefalo, quale principale centro dell'integrazione dell'esperienza emotiva attuale con quelle precedenti.
Così complesse e non sempre, o non ancora, ben chiare implicazioni di ordine psicologico nella percezione del dolore determinano la presenza nei pazienti portatori di neoplasia in stadio avanzato, affetti da malattia dolore, di una serie di alterazioni psichiche, tra le quali predominano l'ansia, la depressione, l'aggressività.
L'ansia, avvertita dal paziente come perdita dell'autocontrollo e delle difese verso il mondo esterno, comporta disturbi psicoemotivi - della memoria, dell'attenzione, della critica, dell'umore - e organici - alterazioni cardiocircolatorie, respiratorie, cenestesiche - che sono responsabili di una serie di paure: del dolore incontrollabile, della mutilazione chirurgica, della morte, della perdita della condizione sociale, del futuro incerto e improgrammabile.
La depressione è un particolare tipo di reazione psichica, anormale per intensità e durata, insorgente nel paziente affetto da una malattia di lunga durata, che comporta perdita della funzionalità fisica, dell'autonomia, dell'immagine corporea, alla quale si associano non infrequentemente problemi economici, difficoltà di reperire aiuto, probabilità di perdere l'attività lavorativa.
L'aggressività, infine, è un fenomeno comportamentale pluriarticolato e complesso, il cui sviluppo sembra si attui attraverso un meccanismo che implica impulsi afferenti al talamo, organizzazione reattiva a livello del sistema limbico e correlazioni con l'area temporale, probabilmente associato a un'aumentata secrezione di noradrenalina (v. Euler, 1956). Nel paziente portatore di neoplasia in fase avanzata il comportamento aggressivo sarebbe indotto e favorito da una particolare disposizione congenita, dal senso di frustrazione per l'inefficacia della terapia e dal risentimento verso la malattia.
Le varie alterazioni psichiche descritte, una volta consolidatesi, possono a loro volta esaltare la sensibilità al dolore fisico, generando così circoli viziosi che aumentano la difficoltà della terapia del dolore. Si comprende quindi come in clinica oncologica sia indispensabile poter disporre di valide informazioni sullo stato psichico del paziente, onde accertare l'esistenza di alterazioni più o meno importanti, fino a quadri francamente paranoidi o schizoidi di non rara osservazione, e procedere così a una più precisa separazione dei problemi di ordine fisiopatologico da quelli di ordine puramente psicologico. All'uopo sono impiegati vari test, tra cui appare di indubbia utilità il cosiddetto MMPI (Minnesota Multiphasic Personality Inventory) in versione tarata per la popolazione italiana: si tratta di un test psicometrico basato su un questionario di 370 domande a ognuna delle quali il soggetto deve rispondere ‛vero' o ‛falso'; le risposte date dal paziente vengono analizzate con una apposita griglia e riportate su un tracciato con tarature differenziate per maschi e femmine.
e) Semeiologia e diagnosi del dolore
Premessa fondamentale a ogni condotta terapeutica è una esatta valutazione diagnostica; nella malattia dolore, in particolare, un attento rilievo semeiologico consente di chiarire l'origine e i meccanismi del dolore nel caso in esame e di apprezzare, con validi controlli statistici, i risultati del trattamento (v. Twycross e Lack, 1983). A tale scopo l'esame del paziente portatore di neoplasia in fase avanzata deve comprendere l'accurato esame obiettivo, in particolare neurologico, la valutazione del grado di evoluzione della malattia, fornito dai dati degli esami strumentali e di laboratorio, e la localizzazione del dolore riferita a uno schema dell'innervazione metamerica del corpo umano. Gli elementi così raccolti consentono di elaborare una mappa del dolore che deve essere integrata da una serie di altre informazioni: modalità di comparsa e ritmo del dolore, sua qualità, esistenza di cause scatenanti o aggravanti, esistenza di fenomeni concomitanti, intensità del dolore.
Riguardo alla modalità di comparsa, si può distinguere il dolore a inizio brusco da quello a inizio lento e graduale, il dolore continuo, il subcontinuo, l'accessionale.
La qualità del dolore, variamente e più o meno esattamente descritta, è espressa con una ricca terminologia: si parla di dolori contusivi, costrittivi, terebranti, folgoranti, urenti, pulsanti, laceranti, ecc. L'uso corretto di tali termini da parte del paziente, in ciò aiutato e guidato dal medico, può rappresentare un valido aiuto nella formulazione diagnostica della natura del dolore. Il dolore da lesione del tessuto nervoso è descritto come pungente, urente o folgorante, e si accompagna a diminuzione o perdita della sensibilità e della funzione motoria. I dolori da lesione di altri tessuti sono invece descritti come gravativi, crampiformi, trafittivi, laceranti, vengono riferiti a determinati metameri corrispondenti al focolaio algogeno e non si accompagnano a deficit neurologici. I dolori misti, dovuti a contemporanea lesione di un certo numero di neuroni e di grosse fibre nervose e di altri tessuti, sono descritti come crampiformi e laceranti e si associano a disestesie e a deficit parziali della sensibilità.
Cause scatenanti o aggravanti il dolore possono essere la posizione, determinati movimenti, l'affaticamento, le condizioni ambientali e atmosferiche; esse sono però scarsamente apprezzabili o non lo sono affatto nei dolori spontanei.
I fenomeni concomitanti al dolore sono individuabili in varie manifestazioni neurovegetative, nell'espressione mimica, nel comportamento reattivo, e vanno accuratamente ricercati e vagliati.
Di particolare importanza è l'apprezzamento dell'intensità del dolore, clinicamente possibile mediante l'impiego dei metodi percettivi basati sulla valutazione soggettiva, da parte del paziente, dell'entità della propria sofferenza. Uno di tali metodi è il McGill pain questionnaire di Melzak, che consente di valutare l'intensità del dolore dal punteggio finale ricavato dai numeri corrispondenti a parole chiave impiegate per descrivere il dolore stesso. Analoghe informazioni quantitative fornisce il metodo dell'analogo visivo di Scott-Huskisson, che consiste nel rappresentare su un segmento di 10 cm l'intensità del dolore, riferendo a zero l'assenza del dolore e a dieci il massimo dolore immaginabile.
Definito il dolore da cui è affetto nei vari aspetti descrittivi, è possibile includere il paziente in studio in una delle tre principali categorie considerate dalla clinica oncologica, secondo cui i pazienti si distinguono in ambulatoriali, ricoverati, domiciliari. Per seguire tutti questi pazienti e raccogliere le informazioni cliniche che li riguardano, nel Servizio di terapia del dolore dell'Istituto Nazionale per lo Studio e la Cura dei Tumori di Milano viene utilizzata una cartella autocompilata giornalmente in cui sono riportati: caratteristiche del dolore, indicate mediante parole chiave corrispondenti a livelli numerici di intensità, quali lieve, molesto, che esaurisce, terribile, micidiale; le ore di sonno; il tempo trascorso in piedi, seduti e coricati; gli effetti collaterali dei farmaci assunti; il livello di attività, o performance status, riferito a una scala da 10 (normalità) a O (totale inabilità o morte). Tutti questi dati, raccolti settimanalmente in una scheda di monitoraggio presso l'ambulatorio di terapia del dolore e combinati con le informazioni sul livello socioculturale e sulle condizioni psicologiche del paziente, permettono un continuo adeguamento dei trattamenti.
4. Terapia del dolore
a) Strategia terapeutica
Il più importante concetto su cui si basa ogni trattamento antalgico è che il controllo del dolore deve essere ottenuto con mezzi che siano il meno possibile tossici e traumatici, di facile attuazione e di basso costo. L'approccio terapeutico deve seguire due direttive principali: 1) essere multimodale, cioè impiegare diverse tecniche contemporaneamente o in sequenza; 2) seguire un programma progressivo in considerazione degli obiettivi che si vogliono raggiungere, delle condizioni del paziente, dei mezzi terapeutici a disposizione. Sulla base di queste premesse è possibile tracciare nel singolo caso un programma terapeutico razionale ed efficace (v. i contributi di Ventafridda, 1981).
Dal punto di vista clinico i pazienti portatori di neoplasie possono essere classificati in tre categorie a seconda dello stadio di avanzamento della malattia: 1) pazienti in procinto di subire o che hanno appena subito un trattamento chirurgico, radiante o chemioterapico; 2) pazienti con tumore in fase avanzata, ma con prognosi di sopravvivenza superiore a tre mesi; 3) pazienti ‛terminali', con evoluzione del tumore rapida e prognosi di sopravvivenza inferiore a tre mesi. I pazienti del primo stadio sono ancora suscettibili di cure causali, che tendono a limitare la crescita del tumore e possono costituire anche un valido trattamento antalgico; per quelli del secondo stadio, nei quali la cura causale non è possibile, è indicato un trattamento dei sintomi, che può talvolta creare le condizioni per tentare ulteriori cure causali; per i pazienti del terzo stadio, infine, l'unico obiettivo realistico è quello di assicurare la migliore qualità di vita possibile, nell'ambito di un'assistenza continua ospedaliera e domiciliare. Sulla scorta di tali acquisizioni la clinica oncologica prevede tre modalità di terapia del dolore: la terapia oncologica specifica, la terapia di lesione, la terapia di modulazione.
b) Terapia oncologica specifica
Questa terapia, distinta in chirurgica, farmacologica e radiante, costituisce il trattamento di prima scelta, comportando diminuzione del dolore conseguentemente alla rimozione della massa neoplastica.
La terapia chirugica può essere radicale, quando consente l'asportazione completa della neoplasia, o palliativa, quando rappresenta un mezzo per limitare le sofferenze causate dal tumore. La chirurgia palliativa è particolarmente efficace nel trattamento dei dolori da ostruzione di un viscere cavo, realizzabile mediante by-pass, stomie addominali, resezione parziale di masse neoplastiche.
La terapia farmacologica prevede l'impiego di chemioterapici e di ormoni. La chemioterapia si effettua con farmaci dotati di attività citotossica e antiproliferativa; è particolarmente indicata nel trattamento dei linfomi e delle leucemie, del cancro della mammella e di quello della prostata, e in generale in tutte le neoplasie caratterizzate da elevati indici mitotici (v. Skipper, 1971). L'ormonoterapia trova il suo impiego nel trattamento dei tumori cosiddetti ormonodipendenti, particolarmente del cancro della mammella, le cui cellule conservino recettori estrogenici ben dimostrabili con le tecniche attuali (v. McGuire e altri, 1975), di quello della prostata e di quello dell'endometrio: al trattamento ormonale di questi tre tipi di neoplasie consegue un risultato antalgico nel 53% dei casi (v. anche chemioterapia antineoplastica; neoplasie: Oncologia umana; sangue: Leucemie).
La radioterapia palliativa si basa principalmente sull'impiego delle particelle ad alta energia, realizzato con la betatronoterapia, o di radioisotopi emettitori di raggi gamma, realizzato con la telecobaltoterapia. La dose di radiazioni necessaria per ottenere un effetto analgesico di solito non supera i 3.000 rad in un piccolo numero di sedute. L'effetto dipende dalla radiosensibilità del tumore e dai trattamenti associati: è quindi variabile da paziente a paziente ed è migliore quando il dolore è causato dalla compressione piuttosto che dall'infiltrazione di strutture nervose. Le migliori indicazioni alla radioterapia palliativa sono rappresentate dalle metastasi ossee, da quelle epidurali, dai tumori del polmone, dai tumori del grosso intestino.
c) Terapia di lesione
Questo tipo di terapia si attua interrompendo il passaggio della sensazione dolorosa mediante intervento lesivo diretto sulle vie nervose. Tuttavia la lesione delle fibre nervose provoca alterazioni di sinapsi, formazione di nuove vie di connessione, ipersensibilità dei neuroni deafferentati, i quali pertanto divengono sede di stimolazione continua con persistenza del dolore: si spiegano così i fenomeni di disestesia, di arto fantasma, di iperpatia e di anestesia dolorosa. Nell'esperimento di Albe-Fessard nel ratto è possibile dimostrare che l'animale, sottoposto a sezione delle radici dorsali del midollo spinale, dopo alcune settimane dall'intervento comincia a rosicchiare l'arto denervato, che scompare gradatamente nel giro di alcuni mesi.
Per questi motivi negli ultimi anni vi è stata una profonda revisione critica degli interventi chirurgici di denervazione nella terapia del dolore, la cui indicazione è attualmente limitata ai pazienti portatori di neoplasia al terzo stadio. La neurochirurgia del dolore si esegue mediante neurotomie, radicotomie, cordotomie; buoni risultati antalgici, soprattutto nelle metastasi ossee da carcinoma della prostata e della mammella, sono stati ottenuti con l'ipofisectomia, il cui meccanismo d'azione tuttavia non è chiaro. Occorre infine ricordare la possibilità di utilizzare nella terapia della malattia dolore dei pazienti portatori di neoplasia al terzo stadio la lobotomia frontale: questo intervento determina l'abolizione della componente emozionale della sensazione dolorosa, che pertanto diviene più tollerabile, e la modificazione della personalità del paziente, che diviene indifferente alla malattia e alla morte (v. Guidetti, 19762).
Oltre a quelli chirurgici esistono trattamenti di neurolesione chimici e termici, il cui impiego nella terapia del dolore è comunque subordinato a particolari requisiti: massima atraumaticità possibile, selettività di azione sulle fibre nervose, incapacità di indurre peggioramento delle condizioni e ulteriori invalidità, possibilità di rigorosa esclusione dei fasci nervosi già deafferentati, facilità di esecuzione, costo contenuto.
I trattamenti chimici si effettuano iniettando con aghi introdotti per via percutanea sostanze come fenolo o etanolo che, provocando denaturazione proteica e vacuolizzazione del nucleo dei neuroni, causano la distruzione delle fibre sensitive e motorie. Questi metodi, tuttavia, sono privi di selettività e la loro indicazione è pertanto limitata solo ad alcuni casi: 1) terapia del dolore addominale da tumore dello stomaco, del grosso intestino, del pancreas, del rene, del fegato, mediante alcolizzazione del ganglio celiaco; il trattamento determina risoluzione del dolore per circa sessanta giorni e provoca, come unico effetto collaterale, una temporanea ipotensione da vasodilatazione splancnica; 2) terapia del dolore trigeminale da neoplasia del cavo orale, del dolore perineale da neoplasia vulvare o rettale, del dolore incoercibile da lesione permanente degli arti inferiori e degli sfinteri, realizzata mediante rizotomie chimiche subaracnoidee; tuttavia il metodo, che trova impiego anche come integrazione della cordotomia percutanea, non è scevro di effetti collaterali indesiderabili, ed è scarsamente impiegato anche per l'imprevedibilità dei risultati, tenuto conto che fibre sensitive decorrono sicuramente anche nelle radici anteriori, mentre con l'inoculazione delle sostanze chimiche si ledono solo le radici posteriori.
Le lesioni termiche si basano sul principio che una temperatura prestabilita e costante provoca una lesione in un'area limitata senza danneggiare le strutture vicine. La lesione viene praticata dopo aver individuato il neurone sensitivo mediante neurostimolazione. Le tecniche di tale tipo più comunemente impiegate sono la cordotomia cervicale percutanea e la termorizotomia del ganglio di Gasser.
1. La cordotomia cervicale percutanea trova impiego nella terapia dei dolori monolaterali toracici da tumore del polmone e della mammella, da invasione del fegato e del rene, da invasioni pelviche da neoplasie dell'utero o da lesioni pelviche da radiazioni, dei dolori dell'arto inferiore da sarcorna o da metastasi; se il dolore non è monolaterale, occorre completare l'intervento con altre tecniche neurolesive o farmacologiche. L'intervento, che si esegue ledendo le fibre della sensibilità dolorifica corrispondenti all'area dolente mediante scariche erogate da un apparecchio di radiofrequenza, provoca perdita della sensibilità termodolorifica dell'emisoma controlaterale, senza perdita del controllo della motilità, e l'analgesia che ne consegue dura da tre mesi a un anno; possono complicare l'intervento una transitoria astenia degli arti omolaterali alla lesione, una disestesia che si manifesta nel 15% dei casi trattati, un deficit funzionale respiratorio da lesione del fascio reticolospinale. Quest'ultima possibilità costituisce un pericolo potenziale che impone di considerare controindicazioni assolute alla cordotomia l'esistenza di un basso tasso di ossigenazione del sangue o di una lesione polmonare controlaterale.
2. La termorizotomia del ganglio di Gasser viene effettuata nella terapia dei dolori trigeminali da neoplasie del cavo orale o della guancia e della nevralgia essenziale del trigemino; è eseguita mediante termolesione prodotta da un elettrodo introdotto per via percutanea nel forame ovale, dopo aver individuato con stimolazioni a bassa frequenza la branca del trigemino che deve essere anestetizzata. L'effetto dell'intervento è l'induzione di analgesia permanente.
d) Terapia di modulazione
Questo tipo di terapia del dolore si attua sia inibendo il passaggio della sensazione dolorosa a livello del sistema nervoso centrale o periferico, senza indurre lesioni neuronali, sia migliorando le condizioni di vita dei pazienti. Le tecniche impiegate sono: blocchi anestetici locali, infusione di oppioidi, elettrostimolazione transcutanea, somministrazione di farmaci analgesici, ipnosi, fisiocinesiterapia.
Blocchi anestetici locali. - Questa tecnica è impiegata nel trattamento del dolore da contratture antalgiche, da alterata statica da forme reumatoidi associate alla neoplasia. Le infiltrazioni di cortisonici e/o di anestetici si effettuano nello spazio epidurale e nel ganglio stellato per ottenere la risoluzione rispettivamente dei dolori sacrolombari e dell'arto superiore, nei cosiddetti punti trigger muscolari e nelle articolazioni per ottenere la remissione dei dolori da contratture muscolari e da flogosi articolari.
Infusione di oppioidi. - Questa tecnica, consistente nella somministrazione di morfina mediante catetere spinale negli spazi peridurale o subaraenoideo, consente di indurre analgesia per circa dodici ore.
Elettrostimolazione transcutanea. - In realtà scarsamente usata in clinica oncologica, questa tecnica è soprattutto indicata nel trattamento della nevralgia posterpetica, del dolore da arto fantasma, delle disestesie postoperatorie.
Somministrazione di farmaci analgesici. - I farmaci in grado di indurre analgesia più comunemente utilizzati nella terapia del dolore appartengono a quattro categorie principali: 1) antinfiammatori non steroidei (FANS); 2) antinfiammatori steroidei (FAS); 3) psicotropi; 4) narcotici. Le modalità del loro impiego sono definite da un protocollo dell'Organizzazione Mondiale della Sanità, che ne indica una somministrazione in sequenza basata sull'intensità del dolore da trattare.
1. FANS. L'azione analgesica di questi medicamenti è sia diretta, basata sull'induzione di iperpolarizzazione nella membrana dei neuroni, con conseguente modulazione della trasmissione dell'impulso algogeno, sia indiretta, legata alle proprietà antiflogistiche fondate sull'inibizione degli enzimi lisosomiali e della sintesi delle prostaglandine e sulla depressione dei livelli di sostanze ossidanti rilasciate nella formazione delle prostaglandine stesse. Il loro effetto nei dolori viscerali è scarso, migliore quello nei dolori provocati da lesioni tendinee, aponeurotiche, del periostio e da metastasi ossee. In generale, tra i numerosi effetti collaterali prodotti da questi farmaci i più temibili sono quelli a carico dell'apparato digerente: gastriti, ulcere gastriche, emorragie. I farmaci dotati di maggiore gastrolesività sono, in ordine decrescente: aspirina, indometacina, derivati dell'acido propionico, difiunisal, paracetamolo.
2. FAS. Essenzialmente rappresentati dai cortisonici, questi farmaci esplicano un'azione antalgica indiretta legata alle loro proprietà antiedemigena e inibente la liberazione delle prostaglandine, cui conseguono a livello delle strutture nervose diminuzione della compressione e inibizione dei fenomeni fiogistici reattivi, entrambi in grado di provocare dolore. Il loro uso è indicato nelle ipercalcemie che si accompagnano ad alcune neoplasie, nelle neuropatie paraneoplastiche, nelle paraplegie iniziali, nei linfedemi, nell'ipertensione endocranica, nella compressione di radici nervose. Controindicazioni assolute all'uso dei cortisonici sono il diabete, il glaucoma, l'ulcera gastroduodenale, la tubercolosi, l'ipertensione arteriosa, le cardiopatie scompensate, le congestioni polmonari, le gravi insufficienze epatiche e renali, il pericolo di emorragie, la presenza di alterazioni psichiche.
3. Psicotropi. L'effetto antalgico di questi farmaci si esplica principalmente mediante la riduzione dei disturbi emozionali che concorrono ad aggravare la sofferenza, e in parte anche mediante meccanismi la cui natura non è del tutto chiara. I principali gruppi di questi farmaci sono i neurolettici, gli ansiolitici, gli antidepressivi. Gli antidepressivi, in particolare, inducono miglioramento del tono dell'umore, aumento delle ore di sonno, facilitazione dei meccanismi endogeni di analgesia; il loro uso è particolarmente indicato nella terapia del dolore da arto fantasma, del dolore da deafferentazione e della nevralgia posterpetica.
4. Narcotici. I narcotici si usano quando gli altri trattamenti non hanno dato risultati soddisfacenti; per il corretto impiego di tali farmaci occorre tener presente che la prevenzione del dolore si ottiene somministrandoli a ore fisse piuttosto che a richiesta del paziente, che è possibile mantenere integro il sensorio del paziente stesso bilanciando la dose in modo da ottenere analgesia senza provocare sonnolenza, che è possibile e preferibile somministrarli per via orale per la maggiore maneggevolezza e la più lunga durata d'azione. I narcotici maggiori, morfina e metadone, devono essere impiegati solo dopo che quelli minori, da soli o in associazione a FANS, si siano dimostrati inefficaci. Il trattamento con narcotici deve essere eseguito con la massima attenzione, in considerazione soprattutto dei fenomeni di tolleranza, accumulo, agonismo parziale. La tolleranza è definibile come la necessità di aumentare progressivamente la dose del farmaco per ottenere lo stesso effetto terapeutico; per accumulo s'intende l'aumento, per somministrazioni successive, della concentrazione plasmatica del farmaco, la cui emivita sia superiore alla durata dell'effetto analgesico; l'agonismo parziale è la caratteristica proprietà di un farmaco, come ad es. codeina e pentazocina, di bloccare l'effetto sui recettori di un agonista puro, come la morfina, insieme col quale siano somministrati in associazione, innescando così una crisi di astinenza. Per questi motivi la morfina somministrata per via orale rappresenta il farmaco preferibile, agonista puro, pratico e maneggevole; i pazienti in cui ne è indicato l'impiego sono i portatori di neoplasie intrattabili, con forti dolori plurifocali, ma senza occlusiofli intestinali o gravi insufficienze epatiche o renali. Gli effetti collaterali indesiderabili conseguenti alla somministrazione di morfina, che vanno accuratamente controllati, consistono essenzialmente in nausea e vomito, sonnolenza, stipsi. Per potenziarne l'azione analgesica, alla somministrazione di morfina si può associare quella di FANS, cortisonici, antidepressivi e tranquillanti per favorire il riposo notturno.
Ipnosi. - Questa pratica deve essere presa in considerazione essenzialmente per il suo aspetto ansiolitico e diversionale, in grado quindi di far diminuire l'attenzione prestata al dolore. Viene eseguita come terapia di gruppo.
Fisiocinesiterapia. - Le varie forme di fisioterapia, cinesiterapia attiva e passiva, associate a massaggi, idroterapia, rieducazione delle stomie, sono impiegate per migliorare le condizioni di vita dei pazienti.
e) Unità di assistenza continua
Nell'ospedale il paziente portatore di neoplasia in fase avanzata soffre non soltanto per la malattia, ma anche perché il personale medico e paramedico presta scarsa attenzione ai suoi bisogni fisici, emotivi, spirituali: è questo l'atteggiamento rinunciatario di una medicina che, aderendo strettamente alla sua metodologia diagnostica e terapeutica, mira a ottenere la guarigione o almeno il prolungamento della vita del malato, apparentemente ignorando che in alcuni casi l'unico obiettivo realisticamente perseguibile è il miglioramento della qualità della vita. Per raggiungere tale obiettivo è necessario superare il modello tradizionale dell'assistenza medica.
Le esperienze degli ultimi quindici anni nei paesi anglosassoni hanno dimostrato che vi è una grande potenzialità di idee e di risorse per venire incontro alle esigenze dei pazienti con neoplasie intrattabili (v. Twycross e Ventafridda, 1980; v. Twycross e Lack, 1983). Il nuovo modello di assistenza che ne risulta si può definire ‛Unità di assistenza continua': si tratta di una struttura aperta in cui il paziente soggiorna per brevi periodi, il cosiddetto Hospice o ‛Unità di cura palliativa', integrata con un ‛Servizio di assistenza domiciliare'. Gli Hospices hanno avuto origine in Irlanda circa cento anni fa, con i ricoveri per i moribondi delle Sisters of charity, e da circa trent' anni vanno diffondendosi nel Canada e negli Stati Uniti.
Il termine di Hospice ha perso la sua connotazione di luogo fisico e viene ora usato per definire un modello di assistenza che è sostanzialmente un programma di supporto per i pazienti e per le loro famiglie, realizzato direttamente o mediante la collaborazione con il medico di famiglia o con altre organizzazioni assistenziali. L'intera famiglia vi viene considerata come un'‛unità' che deve essere assistita, e l'aiuto si prolunga al periodo del lutto che segue la morte del paziente. Viene posta particolare attenzione al controllo dei sintomi e alla preparazione alla morte; sono previsti solo i servizi sanitari essenziali e l'attività di una équipe organizzata interdisciplinarmente, in servizio ventiquattro ore su ventiquattro. Gli obiettivi che si prefigge tale servizio sono: 1) consentire la permanenza in casa del paziente il più a lungo possibile e fare in modo che anche la morte avvenga a domicilio; 2) integrare, piuttosto che duplicare, i servizi sanitari già esistenti nell'ospedale; 3) educare sia gli operatori sanitari, sia le famiglie; 4) assistere l'intera famiglia come unità; 5) aiutare il paziente a migliorare quanto più possibile la qualità della propria vita; 6) diminuire i costi dell'assistenza.
Nell'impossibilità di costituire in breve tempo una simile struttura, in Italia è stata realizzata, presso il Servizio di terapia del dolore dell'Istituto Nazionale per lo Studio e la Cura dei Tumori di Milano, un'unità esterna che si occupa dell'assistenza domiciliare. Questa ‛Unità di assistenza domiciliare' è costituita da medici, infermiere domiciliari, volontari, e segue i pazienti residenti nella città con opportune terapie sintomatiche fino alla morte. I compiti svolti da tale personale sono: 1) adeguare i dosaggi degli analgesici alle esigenze dei singoli casi; 2) offrire la collaborazione alla famiglia per risolvere le difficoltà ambientali e sociali; 3) riferire al medico responsabile ogni cambiamento della situazione del malato; 4) convogliare in una riunione settimanale tutte le esperienze, in modo da coordinare le attività di assistenza sanitaria con quelle di volontariato.
5. Conclusioni
Una vasta e feconda serie di studi sulla fisiopatologia del dolore nell'uomo, pur nella non superata difficoltà di operarne una quantificazione, ha consentito di metterne in evidenza la componente psichica, affettiva ed emotiva che in molti casi rappresenta l'elemento di gran lunga dominante della sofferenza. A questo tipo di dolore rivolge ora la propria attenzione la clinica oncologica, nel tentativo di migliorare le condizioni di vita dei pazienti portatori di neoplasie in fase terminale, fino a un recente passato esclusi da una valida assistenza medica, riservata solo ai casi ancora trattabili.
La moderna terapia del dolore si avvale dei più recenti progressi farmacologici, chirurgici, psico- e fisioterapici per programmare, caso per caso, una strategia terapeutica tesa a rendere tollerabile il periodo terminale del paziente ormai condannato dall'inesorabile progresso del male. Tale strategia terapeutica non può quindi prescindere dalla valutazione degli aspetti psicologici, umani, affettivi, sociali dei singoli pazienti; in breve, deve tendere non soltanto a lenire il dolore, a ottenere un più o meno pronto e duraturo effetto antalgico, ma soprattutto a curare il ‛malato di dolore'. Le prospettive future di questo settore della clinica oncologica sembrano quindi in larga misura dipendenti dal perfezionamento di un'impostazione terapeutica di così nuova concezione nel trattamento del dolore, più ancora che dall'apporto di nuovi contributi tecnici - chirurgici o fisioterapici - o dalla scoperta di nuove molecole chimiche ad azione antalgica.
La terapia del dolore si va affermando, s'è affermata, progredisce col tumultuoso evolversi delle conoscenze medico-biologiche. È al di là delle implicazioni di ordine filosofico, morale e religioso che la sua costruzione teorica comporta, è fuori di dubbio che il suo più grande successo è riuscire a rendere vivibile il periodo terminale di un paziente la cui sofferenza è così intensa e così lesiva della sua personalità da isolarlo completamente dal mondo che lo circonda e renderlo addirittura indifferente alla morte. La terapia del dolore interviene così nella cura delle fasi terminali delle neoplasie, e raggiunge il fine di lenire la sofferenza e forse di infondere nuova speranza, il che, da Ippocrate ai giorni nostri, rimane il più alto ideale della medicina.
(L'organizzazione e l'impianto di questo articolo - al fine di coordinare i contributi dei vari autori - sono stati curati dal prof. Mario Crespi).
bibliografia
Barker, J. L., Physiological roles of peptides in the nervous system, in Peptides in neurobiology (a cura di H. Gainer), New York 1977, pp. 295-343.
Bonica, J. J., Ventafridda, V. (a cura di), Advances in pain research and therapy, vol. II, New York 1979.
Déjerine, J., Roussy, G., Le syndrome thalamique, in ‟Revue neurologique", 1906, XIV, pp. 521-532.
Euler, U.S. von, Noradrenaline, Springfield, Ill., 1956.
Fields, H. L., Basbaum, A. I., Brainstem control of spinal pain transmission neurons, in ‟Annual review of physiology", 1978, XL, pp. 217-248.
Guidetti, B., Terapia chirurgica del dolore, in Enciclopedia medica italiana, vol. V, Firenze 19762, coll. 556-563.
Hughes, J., Isolation of an endogenous compound from the brain with pharmacological properties similar to morphine, in ‟Brain research", 1975, LXXXVIII, pp. 295-308.
Hughes, J., Smith, R.W., Kosterlitz, H. W., Fothergill, L.A., Morgan, B.A., Morris, H.R., Identification of two related pentapeptides from the brain with potent opiate agonistic activity, in ‟Nature", 1975, CCLVIII, pp. 577-579.
Klee, W.A., Endogenous opiate peptides, in Peptides neurobiology (a cura di H. Gainer), New York 1977, pp. 375-396.
Long, D.M., Erickson, D.E., Stimulation of the posterior columns of the spinal cord for relief of intractable pain, in ‟Surgical neurology", 1975, I, pp. 134-141.
McGuire, W. L., Carbone, P. P., Vollmer, E. P., Estrogen receptors in human breast cancer, New York 1975.
Miller, R. J., Enkephalin: a peptide with morphine-like properties, in ‟Trends in neurosciences", 1978, I, pp. 29-31.
Porta, V., Dolore, in Enciclopedia medica italiana, vol. V, Firenze 19762, coll. 517-556.
Ralston, H. J. III, The fine structure of neurons in the dorsal horn of the cat spinal cord, in ‟Journal of comparative neurology", 1968, CXXXII, pp. 275-302.
Rexed, B., The cytoarchitectonic atlas of the sipnal cord of the cat, in ‟Journal of comparative neurology", 1954, C, pp. 297-380.
Skipper, H.E., Cancer chemotherapy is many things. G. H. A. Cloves memorial lecture, in ‟Cancer research", 1971, XXXI, pp. 1173-1180.
Twycross, R.G., Lack, S.A. (a cura di), Symptom control in far advanced cancer: pain relief, London 1983.
Twycross, R.G., Ventafridda, V. (a cura di), The continuing care of terminal cancer patients, Oxford-New York 1980.
Ventafridda, V., Il controllo del dolore, in Manuale di oncologia medica (a cura di G. Bonadonna e G. Robustelli), Milano 1981, pp. 899-908.
Ventafridda, V., Principi di riabilitazione, in Manuale di oncologia medica (a cura di G. Bonadonna e G. Robustelli), Milano 1981, pp. 909-914.
Ventafridda, V., La cura del paziente terminale, in Manuale di oncologia medica (a cura di G. Bonadonna e G. Robustelli), Milano 1981, pp. 917-921.
Yaksh, T.L., Rudy, T.A., Narcotic analgesics: CNS sites and mechanisms of action as revealed by intracerebral injection techniques, in ‟Pain", 1978, IV, pp. 299-359.
© Istituto della Enciclopedia Italiana - Riproduzione riservata


