Protisti
Enciclopedia della Scienza e della Tecnica (2007)
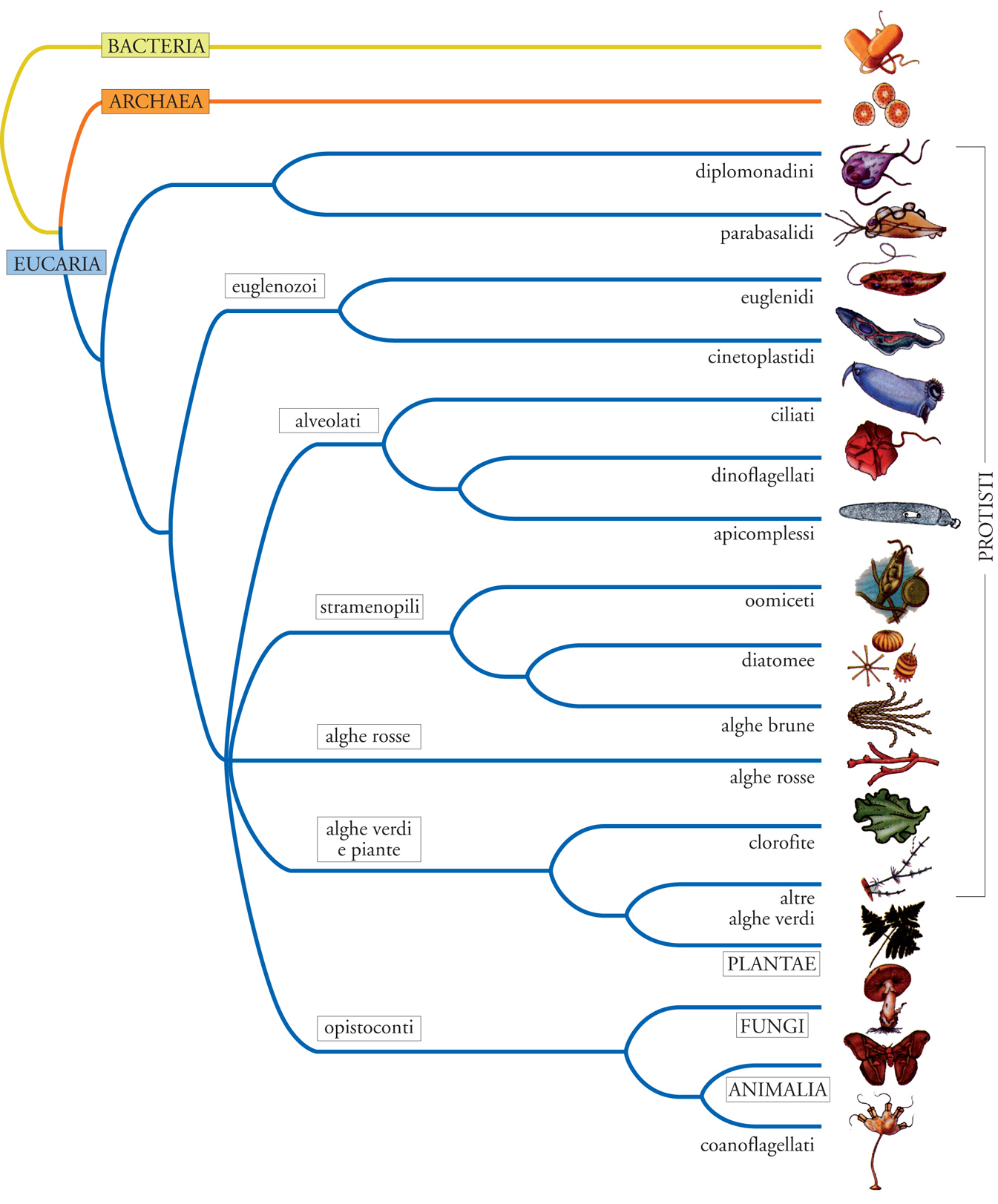
Protisti
Si identificano come protisti tutti quegli organismi eucarioti (formati, cioè, da cellule con cromosomi collocati all'interno di un nucleo) che possiedono un'organizzazione corporea fondamentalmente unicellulare, o comunque evoluta a un livello di complessità non equiparabile a quello di tessuto e organo che contraddistingue ogni rappresentante dei tre regni degli eucarioti pluricellulari (Plantae, Fungi e Animalia). Tra i protisti si ritrovano tutte le prime forme di vita eucariotica, così come tutti i capostipiti evolutivi della biodiversità pluricellulare. Da un punto di vista filogenetico i protisti comunque non costituiscono un regno protista a sé stante, perché non rappresentano un'entità monofiletica in cui è possibile identificare un progenitore e i suoi discendenti evolutivi, sulla base di comuni caratteri derivati (o sinapomorfie). Essi piuttosto rappresentano una vasta ed eterogenea confederazione parafiletica di numerosi gruppi, di rango tassonomico equiparabile a phylum, che hanno avuto modo di separarsi evolutivamente, in un arco di tempo di almeno un miliardo di anni, in modo assai più profondo di quanto sono tra loro separati piante, funghi e animali.
Una consuetudine originata da un inquadramento tassonomico della biodiversità prioritariamente basato su una dicotomia tra i regni vegetale e animale vuole che una parte di questi gruppi, in quanto fotosintetici (o autotrofi), sia classificata come protofite, o alghe, e inclusa nel regno vegetale, e una parte come Protozoi, in quanto fagotrofici (o eterotrofi), e inclusa nel regno animale. A questo riguardo è necessario puntualizzare che i taxa Algae e Protozoa non possono, come tali, avere un riconoscimento scientifico formale in un inquadramento sistematico dei protisti che sia mirato a rifletterne fedelmente la storia evolutiva. L'alternativa tra autotrofia ed eterotrofia, che interessa le piante e gli animali, non interessa assolutamente i protisti. Non solo vari gruppi filogeneticamente omogenei (per es., i dinoflagellati) comprendono sia specie autotrofe che eterotrofe, ma addirittura uno stesso organismo (per es., Euglena) può indifferentemente comportarsi da autotrofo, o eterotrofo, secondo che viva alla luce, o al buio.
La suddivisione formale dei protisti tra alghe e Protozoi ha tuttavia mantenuto una sua indubbia utilità pratica in campo sia didattico, sia divulgativo, ed è stato ritenuto opportuno rispettarla anche in questo contesto. La trattazione sarà comunque circoscritta ai protisti Protozoi, i quali rappresentati dai gruppi (con valore tassonomico di phylum) qui di seguito riportati in ordine alfabetico e con un'indicazione del numero approssimativo di specie viventi che li rappresentano.
Actinopoda (acantari, eliozoi e radiolari; 500 specie), Amoebozoa (rizopodi, 500 specie); Apicomplexa (gregarine, emosporidi, coccidi e piroplasmi; 5000 specie); Choanoflagellata (150 specie); Ciliophora (ciliati; 8000 specie); Dinozoa (dinoflagellati; 2000 specie); Euglenozoa (euglene, leishmanie e tripanosomi; 2000 specie); Eumycetozoa (amebe aggreganti; 700 specie); Foraminifera (6000 specie); Hemimastigophora (1 specie); Heterolobosea (schizopirenidi; 50 specie); Microsporidia (800 specie); Myxospora (mixozoi; 1200 specie); Opalinida (400 specie); Parabasalia (tricomonadini e ipermastigini; 500 specie); Pseudociliata (3 specie); Xenophyophorea (50 specie).
Euglenozoa
Sotto la denominazione di Euglenozoa si unificano gli euglenidi veri e propri e i cinetoplastidi (tripanosomi e leishmanie) in una stessa linea filogenetica; si tratta presumibilmente di uno dei più antichi tra i gruppi di protisti. Tutti possiedono mitocondri con creste discoidali (in pratica, a forma di racchetta da ping-pong), che ne giustificano anche la denominazione, insieme agli Heterolobosea, di Discicristata. Analisi genetiche molecolari inoltre suggeriscono una loro discendenza da un comune progenitore dotato di alghe endosimbionti fotosintetiche, la cui presenza sarebbe oggi rappresentata, negli euglenidi, dai plastidi (delimitati da tre membrane, invece che da due come nelle piante) e, nei cinetoplastidi, da specifiche sequenze genomiche. Negli euglenidi coesistono le tre divisioni di ordine trofico che separano animali (fagotrofia), piante terrestri (fototrofia) e funghi (saprotrofia). Ne fanno infatti parte sia specie fotoautotrofe verdi, sia specie incolori ed eterotrofe, prive di cloroplasti e capaci di assumere nutrienti organici per saprotrofia e materiale particolato per fagocitosi. Varie specie possono inoltre passare da un tipo di metabolismo all'altro secondo le condizioni ambientali in cui vengono a trovarsi.
Questa plasticità metabolica favorisce molto le esplosioni demografiche ('fioriture') degli euglenidi in acque ricche di materiale organico in decomposizione, in occasione delle quali si possono raggiungere concentrazioni di oltre un milione di cellule/ml. Gli euglenidi tipicamente possiedono due flagelli impiantati in una profonda depressione della superficie cellulare anteriore (tasca flagellare, o reservoir), ma in Euglena uno dei due si riduce a un corto moncone, mentre l'altro si ricopre dei sottili filamenti noti come 'mastigonemi', che ne aumentano l'efficienza propulsiva. In corrispondenza della tasca flagellare si svuota anche un grosso vacuolo contrattile (pusula) e risiedono due organuli con funzione di fotorecettori, rappresentati da un 'corpo paraflagellare' costituito da un cristallo proteico localizzato in un rigonfiamento della radice flagellare e da una 'macchia ocellare' (di colore rosso-arancio per il suo contenuto di carotenoidi).
I cinetoplastidi sono così chiamati in relazione a un loro specifico organulo, il cinetoplasto, portato all'interno di un unico voluminoso mitocondrio. Il cinetoplasto contiene un suo DNA (noto come kDNA) organizzato a formare un complesso di molecole circolari tutte tra loro connesse, ma distinte per dimensioni, numero e attività. Per quanto i cinetoplastidi comprendano anche numerose specie di bodonidi a vita libera, la loro notorietà è preminentemente legata all'interesse medico, veterinario e agronomico che rivestono le specie di Trypanosoma e Leishmania, parassiti patogeni di uomo, animali domestici e piante. I cicli biologici di queste specie possono compiersi in uno o due ospiti e comportano il differenziamento di vari stadi (noti come 'promastigote', 'opistomastigote', 'coanomastigote', 'amastigote' e 'tripomastigote') tra loro distinti sia per la posizione della radice del flagello rispetto all'estremità anteriore della cellula, sia per l'assenza/presenza di una connessione citoplasmatica (membrana ondulante) tra la superficie cellulare e la porzione extracellulare del flagello. Nel caso delle leishmanie, diffuse nelle aree sia tropicali sia subtropicali, l'infezione è causata dalle punture dei flebotomi e l'uomo contrae una varietà di malattie collettivamente note come 'leishmaniosi' cutanee e viscerali. La più grave è il kala-azar (oltre un milione di persone infette e un migliaio di decessi ogni anno) generata dall'invasione di fegato, milza e midollo spinale da parte dello stadio amastigote (di dimensioni di soli 2÷3 μm) di Leishmania donovani.
Nel caso dei tripanosomi, invece, i vettori variano a seconda che si tratti di Trypanosoma cruzi dell'America Centrale o di T. brucei della fascia equatoriale dell'Africa. Nel primo caso, la trasmissione del parassita è causata da varie specie di Triatoma e di altri eterotteri reduvidi (complessivamente noti come kissing bugs, poiché solitamente pungono chi dorme nelle aree molto vascolarizzate circostanti la bocca); nel secondo, da due specie di Glossina (G. morsitans e G. palpalis) note come 'mosca tse-tse'. Mentre T. cruzi è comunemente denominato 'tripanosoma stercorario', perché fuoriesce dalle cimici con le feci e penetra poi nel sangue della vittima attraversandone le mucose, T. brucei è noto come 'tripanosoma salivare', poiché passa dall'intestino medio nel lume delle ghiandole salivari per essere iniettato nel sangue della vittima dalla puntura della mosca tse-tse. Annualmente nell'America Centrale e Meridionale, T. cruzi infetta (ed è causa del morbo di Chagas) da 15 a 20 milioni di persone, delle quali varie migliaia muoiono per danni irreparabili, soprattutto a livello dei tessuti muscolari (pareti del cuore comprese) e nervosi. Altrettanto alta è in Africa l'incidenza della malattia del sonno (quasi 100.000 decessi l'anno) generata da T. brucei al momento in cui questo parassita passa dal sangue nel sistema linfatico e nel liquido cerebrospinale.
I tripanosomi hanno sviluppato un meccanismo molto efficace per eludere la risposta immunitaria dell'ospite. Una volta penetrati nel sistema circolatorio, la loro superficie cellulare si riveste di una glicoproteina molto antigenica che ha la proprietà di poter essere sintetizzata in numerose forme diverse, ognuna specificata da una sua sequenza genica. Di queste varie forme, i tripanosomi a ogni modo ne esprimono sulla superficie cellulare solo una alla volta e soltanto per alcuni giorni, per poi procedere alla sintesi ed esposizione di una nuova. In tal modo gli anticorpi prodotti dall'ospite contro una determinata forma, al momento di entrare in circolazione trovano un bersaglio sistematicamente diverso; così la loro efficacia è pressoché nulla. In pratica i tripanosomi riescono a stare sempre un passo più avanti della risposta immunitaria dell'ospite.
Diplomonadina

I diplomonadini debbono la loro denominazione all'assetto simmetrico bilaterale che caratterizza la struttura del corpo cellulare delle specie filogeneticamente più moderne, nelle quali due complessi nucleo-flagellari (o cariomastigonti), ognuno tipicamente formato da un nucleo e quattro flagelli, si dispongono in modo tale da apparire uno l'immagine speculare dell'altro. Questi due complessi hanno verosimilmente avuto origine da una duplicazione di un complesso originariamente unico, che invece caratterizza la struttura delle specie più antiche, note come 'enteromonadini'. I diplomonadini vivono prevalentemente nell'intestino degli animali, mentre le poche specie a vita libera prediligono ambienti con forte inquinamento organico e poveri di ossigeno. Le forme intestinali si comportano in genere da commensali innocui; tuttavia, Giardia intestinalis (fig. 6) è patogena per l'uomo, mentre Hexamita salmonis e H. meleagridis sono causa di forti morie negli allevamenti, rispettivamente, di trote e salmoni e di galliformi (tacchini, in particolare). Un importante aspetto morfologico di queste forme, che ha indotto a ipotizzare una posizione filogenetica dei diplomonadini addirittura alla radice dell'evoluzione degli eucarioti, è l'assenza nel loro citoplasma di un corpo di Golgi, di lisosomi, reticolo endoplasmico liscio e, soprattutto, mitocondri. Analisi molecolari tuttavia non avvalorano assolutamente questa posizione dei diplomonadini e si deve pertanto concludere che la mancanza di mitocondri e delle altre strutture citoplasmatiche abbia un'origine secondaria.
Parabasalia
Come nel caso dei diplomonadini, anche questo raggruppamento racchiude flagellati endosimbionti anaerobi privi di mitocondri, funzionalmente sostituiti da idrogenosomi che, trasformando il piruvato in acetato, producono sia energia per reazioni enzimatiche di fosforilazione sia idrogeno molecolare. I parabasalidi debbono il nome a una struttura citoscheletrica, nota come 'corpo parabasale', formata da un complesso di fibrille con striatura trasversale periodica che connettono fisicamente il corpo di Golgi alla radice (corpuscoli basali) dei flagelli. Associate a questo corpo parabasale ci sono usualmente altre strutture citoscheletriche, sia di natura fibrillare e striate (pelta e atrattoforo) sia microtubulare (axostilo), funzionalmente deputate a fornire una maggiore stabilità strutturale alla cellula durante il movimento. Tra i parabasalidi si distinguono i tricomonadini ‒ dotati di 4-6 flagelli di cui uno (flagello ricorrente) è di norma diretto posteriormente e fisicamente connesso alla superficie cellulare mediante una membrana ondulante ‒ dagli ipermastigini caratterizzati da un imponente complesso di centinaia di flagelli cui spesso si associano anche spirochete epibionti che si muovono coordinatamente con i flagelli stessi. I primi vivono da parassiti nell'intestino e nell'apparato urogenitale di un ampio spettro di animali, e annoverano anche alcune specie patogene per l'uomo (Trichomonas vaginalis) e per gli animali domestici (come Histomonas meleagridis nei polli). I secondi sono endosimbionti obbligati nell'intestino degli Insetti xilofagi (termiti e blatte in particolare), ai quali forniscono tutte le cellulasi necessarie per digerire le particelle di legno inglobate da questi per fagocitosi. Tale rapporto di simbiosi è verosimilmente molto antico, al punto che i processi sessuali degli ipermastigini sono ciclicamente indotti dall'innalzamento del livello di ecdisone nell'insetto ospite al momento di entrare in muta.
Choanoflagellata
I coanoflagellati sono rappresentati da specie, solitarie e coloniali, tra le più piccole di tutti i protisti (in media, 10 μm), distribuite nell'acqua dolce come nel mare, dove in genere si presentano dotate di una delicata struttura protettiva (lorica) fatta di bastoncelli silicei (coste) di vario aspetto. Il loro unico flagello è tipicamente circondato da un collaretto fatto da una ventina di proiezioni digitiformi (dall'apparenza di microvilli), con funzioni di intrappolare e dirigere le particelle di cibo (batteri soprattutto) verso la base del collaretto stesso dove si apre un citostoma. Poiché questo collaretto è strutturalmente e funzionalmente equivalente (omologo) a quello dei coanociti che tappezzano lo spongocele dei poriferi, si è da qualche tempo fatta strada l'ipotesi di una stretta parentela evolutiva dei coanoflagellati con i poriferi stessi. Attualmente questa ipotesi non soltanto trova ampio conforto anche in analisi genetiche molecolari, ma si va sempre più accreditando l'ulteriore ipotesi che i coanoflagellati siano addirittura i progenitori evolutivi dell'intero regno degli Animalia (come richiamato anche nell'albero filogenetico della fig. 2). È a ogni modo necessario osservare che la mancanza di ogni chiara correlazione filogenetica tra coanoflagellati e qualsiasi altro gruppo di Protozoi rende altrettanto valida l'ipotesi che i coanoflagellati siano in realtà dei poriferi che sono andati incontro secondariamente a fenomeni di quasi completa disgregazione e dissociazione cellulare.
Opalinida
Le opaline vivono da endosimbionti nell'intestino posteriore dei Vertebrati a sangue freddo e sono particolarmente comuni negli anuri. L'intera loro superficie cellulare ricoperta da ciglia, ordinatamente disposte in fila e sincronizzate nel battito, unitamente alla presenza nel loro citoplasma di più di un nucleo (da due fino anche a centinaia), hanno per lungo tempo fatto pensare a una loro correlazione filogenetica con i ciliati. Analisi ultrastrutturali e molecolari hanno tuttavia definitivamente chiarito che le loro parentele evolutive non sono con i ciliati, ma ricadono nell'ambito dei protisti flagellati, anche se rimane ancora molto incerta l'identificazione di quali siano i gruppi più direttamente coinvolti. Le opaline presentano cicli biologici piuttosto complessi, dei quali il più conosciuto è senz'altro quello di Opalina ranarum, la cui fase asessuale si compie nell'intestino retto delle rane adulte, mentre quella sessuale interessa i girini.
Pseudociliata
Gli pseudociliati sono rappresentati dall'unico genere plurinucleato Stephanopogon, abitante dell'ambiente marino interstiziale dove si nutre principalmente di diatomee. Per lungo tempo ritenuto un ciliato, Stephanopogon ha poi rivelato di non possedere alcuno dei caratteri distintivi dei ciliati, cioè dimorfismo nucleare, complesso di alveoli corticali, infraciliatura e coniugazione. I suoi nuclei sono morfologicamente uguali, la superficie cellulare è formata dalla sola membrana plasmatica sostenuta da una rete microtubulare e le ciglia si muovono con movimento ondulatorio come flagelli e senza coordinamento metacronico. La riproduzione infine avviene per divisioni multiple spesso associate a stadi di regressione ciliare interpretati come stadi di incistamento.
Hemimastigophora
Questo gruppo è noto solo dal 1998 ed è rappresentato da una singola specie di piccole dimensioni (10÷15 μm), Hemimastix amphikineta, isolata da campioni di suolo umido raccolti da Paesi di origine gondwaniana (Australia e Cile). H. amphikineta è uninucleata, eterotrofa per quanto priva di un citostoma permanente, e si muove 'camminando' sui detriti del suolo per mezzo di una ventina di lunghe ciglia ordinate in due linee longitudinali a decorso leggermente a spirale, che conferiscono all'organismo un aspetto simile a quello di un ciliato.
Amoebozoa
Gli amebozoi (o rizopodi) costituiscono un eterogeneo raggruppamento di amebe asessuate, anche di notevoli dimensioni (oltre 2÷3 mm), che hanno in comune unicamente la capacità di formare pseudopodi di aspetto tubuliforme o lobato (lobopodi), o filiforme e ramificato (filopodi), a ogni modo sempre privi di un asse scheletrico interno di natura microtubulare e mai anastomizzati tra loro. Essendo privi di un citostoma, gli amebozoi si nutrono per fagocitosi e pinocitosi attraverso la superficie cellulare formata dalla sola membrana plasmatica. Questa può presentarsi protetta esternamente da una teca (o guscio) di varia forma (sferica, discoidale, piriforme) e natura (sia organica sia silicea, con o senza associazione di materiale esogeno), oppure essere del tutto priva di ogni protezione meccanica per l'intera sua estensione. Il primo caso è rappresentato dai Testacealobosea (o 'amebe col guscio', come Arcella e Difflugia), di piccole dimensioni (non oltre 100 μm) e frequenti in particolare nel terreno umido, spesso in associazione a muschi. Il secondo è invece rappresentato dai Gymnamoebia (o 'amebe nude') con specie endosimbionti commensali in una vasta gamma di altri organismi, uomo compreso, oltre che a vita libera in acque stagnanti sia ossigenate (Amoeba, Chaos, Mayorella) sia anossiche (Pelomyxa, ameba questa che, per il livello primitivo della sua organizzazione strutturale, viene anche considerata come unico rappresentante di una linea filogenetica a sé stante, denominata Pelobiontida).
Tra le specie commensali dell'uomo, Entamoeba hystolitica si comporta da vero parassita patogeno ed è, infatti, responsabile della dissenteria amebica che ogni anno interessa alcuni milioni di persone con numerosi decessi. Nell'intestino crasso, dove E. hystolitica si localizza una volta ingerita in forma di cisti associate a cibo e acqua inquinati, si sviluppano usualmente due forme distinte per dimensioni tra 'minuta' (12÷15 μm) e 'magna' (20÷40 μm). Veramente dannosa è la forma magna, poiché produce enzimi idrolitici che creano ulcere nella parete intestinale, favorendone l'ingresso nel flusso sanguigno e la successiva penetrazione in vari organi come fegato e polmoni; la forma minuta si limita a nutrirsi di polisaccaridi e secrezioni mucose della parete intestinale.
Heterolobosea
Gli eterolobosi (noti anche come 'ameboflagellati') si identificano con alcune specie di amebe uninucleate (Wahlkampfia, Naegleria) le cui correlazioni filogenetiche sono da ricercarsi con gruppi di flagellati più che di altre amebe. Anche gli pseudopodi molto 'eruttivi', che contraddistinguono queste specie durante il loro stadio ameboide, normalmente assunto nel terreno umido ricco di batteri, sono ritenuti non omologhi agli pseudopodi degli Amoebozoa. In ambienti acquatici con scarse risorse alimentari, lo stadio ameboide ha vita breve e in sua vece si sviluppano stadi sia di cisti sia bi- o tetraflagellati, capaci di disperdersi più rapidamente alla ricerca di nuove risorse alimentari. Nel caso di N. fowleri, una specie termofilica, questi stadi flagellati sono pericolosi per l'uomo, in quanto possono penetrare accidentalmente nelle coane nasali (come può avvenire durante un bagno in piscine all'aperto), localizzarsi a livello encefalico e provocare meningoencefaliti anche letali.
Eumycetozoa
Gli eumicetozoi comprendono amebe legate ad habitat terrestri ricchi di materiali in decomposizione e vegetazione morta, in parte caratterizzate da cicli biologici che comportano stadi multinucleati, e in parte da cicli biologici con stadi multicellulari. Gli stadi multinucleati sono distintivi delle specie collettivamente note come 'muffe acellulari del suolo'; la loro organizzazione riflette quella di un plasmodio vero e proprio formato da un unico corpo citoplasmatico condiviso da migliaia (fino a milioni) di nuclei e delimitato esternamente dalla sola membrana plasmatica (corpo che può giungere a un peso anche di decine di grammi ed espandersi su una superficie di oltre 1 m2). Gli stadi multicellulari sono invece distintivi delle specie collettivamente note come 'muffe cellulari del suolo'; la loro organizzazione è considerata come uno pseudoplasmodio, poiché le cellule che lo compongono evitano ogni mutua fusione della loro membrana plasmatica e mantengono ognuna la propria individualità.
Nelle specie plasmodiali (Physarum polycephalum) lo stadio di plasmodio si fa carico dell'intera fase trofica (di accrescimento) del ciclo e si protrae finché l'ambiente offre adeguate risorse alimentari (batteri soprattutto), oltre a sufficiente stabilità di umidità e pH. Al sopraggiungere di condizioni sfavorevoli, il plasmodio può intraprendere due diverse trasformazioni, delle quali una è rappresentata da una massa irregolare e frammentata (sclerozio) che si protegge dall'ambiente ispessendo e sclerotizzando la sua superficie cellulare, ma che rimane in ogni modo capace di riconvertirsi rapidamente a plasmodio trofico nel caso l'ambiente divenga nuovamente favorevole. L'altra trasformazione è invece rappresentata da un corpo fruttifero dall'aspetto molto reticolato, da cui si sviluppano steli tozzi e ramificati (sporangiofori) che terminano apicalmente con ammassi di spore (sporangi), ognuna contenente un nucleo aploide derivato dalla meiosi dei nuclei diploidi del plasmodio. A 'maturazione' del corpo fruttifero, queste spore si disperdono nell'ambiente e liberano singole amebe flagellate capaci di dividersi e disperdersi ulteriormente prima di differenziarsi in gameti. Dall'unione due a due di questi gameti si formano zigoti da cui hanno origine i nuovi plasmodi, con un processo che prevede ripetute divisioni nucleari senza alcuna divisione citoplasmatica.
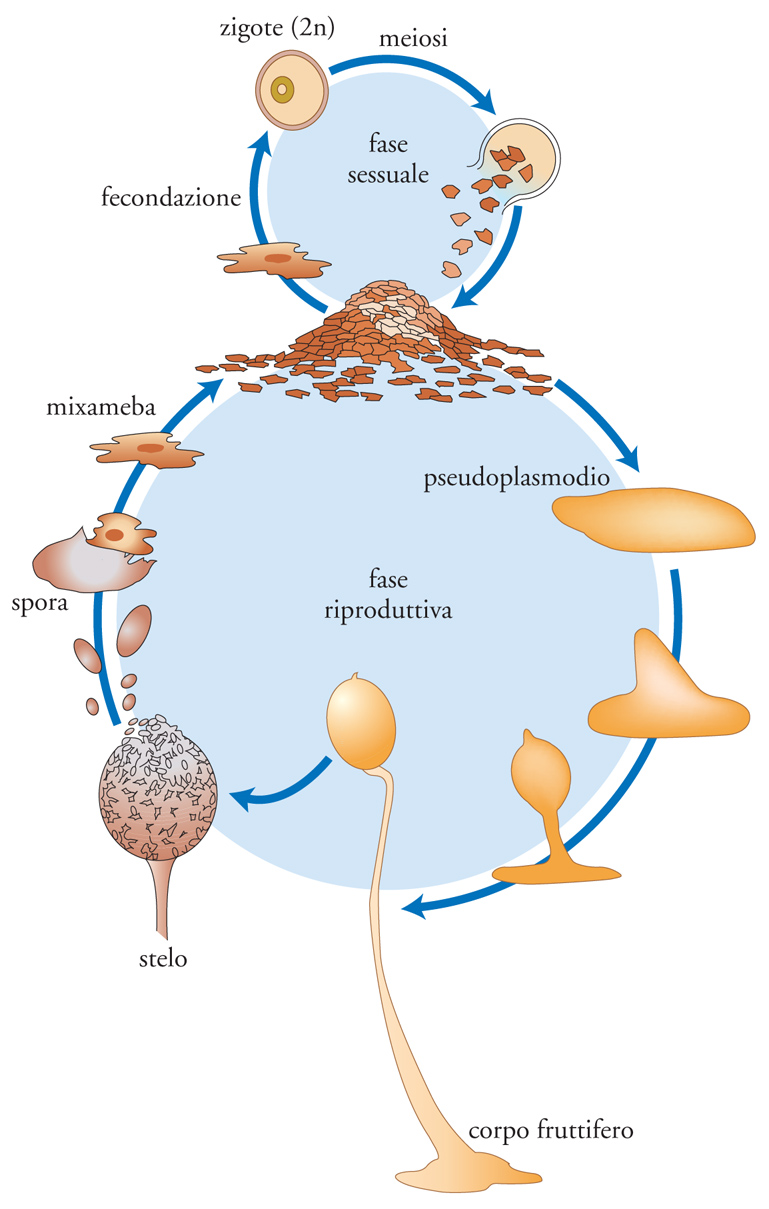
Nelle specie pseudoplasmodiali (Dictyostelium discoideum), diversamente da quelle plasmodiali, sono singole cellule ameboidi (mixamebe) a farsi carico della fase trofica del ciclo, fagocitando batteri e altro materiale particellato per moltiplicarsi poi per divisione mitotica (fig. 7). Al deteriorarsi delle risorse ambientali, queste mixamebe rispondono con la secrezione di AMP ciclico (la stessa molecola ben conosciuta per la sua attività più generale come 'secondo messaggero' citoplasmatico) che, agendo da segnale chemotattico, ne stimola l'aggregazione a formare uno pseudoplasmodio. Una volta acquisita compattezza, questo pseudoplasmodio inizia a scivolare (come una lumaca) sul substrato (grazie anche alla secrezione di una pellicola extracellulare mucopolisaccaridica) finché, dopo alcune ore di 'migrazione', si arresta per modificarsi in corpo fruttifero, le cui spore si disperdono da un unico sporangio sferico portato alla sommità di un lungo sottile stelo. Le mixamebe che fuoriescono iniziano una nuova fase trofica se l'ambiente è compatibile con le loro necessità energetiche; altrimenti esse si differenziano in gameti che si fondono in complessi multipli, al cui interno si forma in ogni caso un unico zigote capace di produrre a sua volta mixamebe con nucleo aploide al termine di una serie di divisioni nucleari.
Actinopoda
Gli attinopodi comprendono radiolari, acantari e eliozoi, tutti dotati di pseudopodi rigidi a forma di raggio (assopodi), formati da un asse interno di microtubuli ricoperto da una pellicola di citoplasma che scorre sia dal centro alla periferia per trasportare sostanze deputate alla cattura di particelle alimentari, sia dalla periferia al centro, per trasportare le particelle catturate e inglobate in vacuoli digerenti. Il loro corpo cellulare inoltre condivide un'organizzazione a simmetria sferica e si presenta generalmente protetto da uno scheletro di natura minerale.
I radiolari sono esclusivamente marini planctonici, distribuiti fino a profondità di 4000÷5000 m, con specie anche coloniali (che possono raggiungere dimensioni di 2 cm), spesso dotati di vivaci pigmentazioni dovute alla presenza di dinoflagellati endosimbionti. La loro struttura è caratterizzata da una capsula centrale chitinosa, pervia per la presenza di numerosi micropori, o (nei feodari) di tre principali aperture (un astropilo e due parapili), che fa da base di impianto agli assopodi e separa il citoplasma in una regione interna intracapsulare e una esterna extracapsulare delimitata dalla sola membrana plasmatica. Nella prima sono presenti uno o più nuclei con materiali di riserva e cristalli proteici molto rifrangenti; la seconda (calimna) è ricca di vacuoli, sia digestivi sia con funzione idrostatica contenenti lipidi e/o elementi allo stato gassoso, e contiene (sempre nei feodari) un ammasso granulare giallo-bruno di materiali di rifiuto molto appariscente (feodio). La struttura dei radiolari in generale si complica molto per la produzione di uno scheletro interno di opale le cui spicole possono saldarsi tra loro in un reticolo sferico, o in altre architetture dall'aspetto elegante quanto bizzarro. Questi scheletri concorrono alla formazione dei sedimenti marini a grandi profondità (noti come 'fanghi a radiolari'), dove rimangono pressoché integri nonostante le elevatissime pressioni poiché l'opale che li compone non si dissolve.
Gli acantari, marini e planctonici come i radiolari, si distinguono per la produzione di uno scheletro di celestite (solfato di stronzio) costituito di 20 spine radiali che si riuniscono al centro della cellula ed emergono in punti ben precisi della superficie cellulare con la porzione apicale ricoperta da ispessimenti del citoplasma periferico extracapsulare ricchi di fasci di mionemi contrattili noti come 'miofrischi'. La disposizione spaziale di queste spine segue un preciso disegno geometrico (o legge di Muller) basato su un'analogia tra la superficie cellulare e quella terrestre. Due quartetti di spine, infatti, emergono a 60° N e 60° S, due a 30° N e 30° S e uno a 0°, e pertanto si identificano, rispettivamente come 'polari', 'tropicali' ed 'equatoriali'. Inoltre, poiché ciascun quartetto è girato di 45° rispetto a quello vicino, le spine polari ed equatoriali emergono a longitudine 0°, 90°, 180° e 270°, e quelle tropicali a longitudine 45°, 135°, 225° e 315°.
Gli eliozoi abitano anche gli ambienti d'acqua dolce oltre che marini. Il loro corpo sferico è spesso peduncolato per aderire al substrato e lo scheletro ha origine da spicole di natura esogena che si cementano tra loro e con vario materiale organico corpuscolato. Il citoplasma più periferico è ricco di vacuoli sia alimentari sia pulsanti, mentre quello più interno alloggia (nelle forme più evolute) un centro organizzatore di microtubuli (noto come 'centroplasto') in stretta associazione con il nucleo. Da questo centro si irradiano in ogni direzione (come raggi del sole) gli assopodi, coinvolti nella cattura del cibo costituito anche da piccoli Metazoi come rotiferi e copepodi, che sono paralizzati con la scarica di estrusomi. Il loro asse interno è costituito da un nutrito fascio di microtubuli disposti secondo un preciso ordine geometrico, che in genere comporta la ripetizione di un'unità di base formata da tre microtubuli disposti a triangolo equilatero, o da sei disposti a esagono. Alla mancanza di risorse alimentari, le forme delle pozze d'acqua temporanee rispondono con la produzione di zoospore flagellate destinate a disperdersi in nuovi ambienti, o entrando in autogamia.
Foraminifera
Sommando le circa 30.000 specie estinte (descritte sulla base di reperti fossili dei loro gusci) alle circa 8000 specie viventi, i foraminiferi costituiscono il gruppo di protisti più numeroso, oltre che uno tra i più rappresentati nel plancton di tutti i mari. I loro pseudopodi sono tipicamente filiformi e connessi uno con l'altro mediante una rete di anastomosi che ne giustifica la denominazione di 'reticolopodi'. Oltre a facilitare il galleggiamento e catturare il cibo, essi sono coinvolti nella costruzione di un guscio (fino anche a 14 cm di diametro nei nummuliti fossili del Cambriano inferiore) che comporta comunque la sintesi di un'iniziale matrice di mucopolissaccaridi. Questa matrice resta tale nei gusci di natura organica che pertanto rimangono assai flessibili e plastici, mentre nei gusci calcarei (largamente i più rappresentati) si arricchisce di cristalli di calcite che conferiscono al guscio aspetti diversi (porcellanato, ialino, o microgranulare) in accordo al modo di aggregazione.
I gusci calcarei degli individui morti si depositano sul fondo marino in quantità enormi a profondità che comunque non superano i 3000÷4000 m (oltre le quali la pressione idrostatica rende solubile il carbonato di calcio), e vanno a costituire i cosiddetti 'fanghi a foraminiferi' spessi fino a decine di metri e utilizzabili anche come materiale edile (per es., parte dei blocchi usati nella costruzione delle piramidi egizie sono calcite originata da gusci di foraminiferi). Questi gusci hanno sempre un'unica apertura per la fuoriuscita dei reticolopodi, e racchiudono una sola camera nelle forme più antiche (pertanto definite 'monotalamiche'), e più camere tra loro comunicanti per mezzo di foramina (da cui trae origine il nome di foraminiferi) in quelle più recenti (politalamiche). La disposizione geometrica di queste camere varia significativamente anche a livello interspecifico (potendo essere in fila, a spirale piana o elicoidale, ad anelli concentrici e così via), ed è conseguita per apposizione progressiva di ciascuna camera a partire da una iniziale (nota come proloculum), che negli stadi sessuali (aploidi) ha usualmente dimensioni maggiori rispetto a quelli asessuali (diploidi).
Xenophyophorea
Gli xenofiofori sono un gruppo di una cinquantina di specie di Protozoi 'giganti' (tra 0,2 e 7 cm di diametro) esclusivamente marini e bentonici di profondità, con corpo cellulare costituito da un plasmodio multinucleato racchiuso in una specie di conchiglia dall'aspetto tipicamente poroso e bitorzoluto, nonché ricco di cristalli citoplasmatici di barite con funzioni ignote. Questa conchiglia è costituita da materiale organico trasparente, che porta associate sia microsfere 'fecali' (stercomi) di natura endogena, verosimilmente originate da aggregazione di materiale non digerito di origine batterica, sia particelle esogene costituite da detrito di altri organismi (come spicole di spugne e conchiglie di foraminiferi e radiolari) e denominate 'xenofi' (da cui trae appunto origine il termine Xenophiophorea, 'portatori di particelle straniere'). Per quanto questi organismi siano stati ripetutamente rinvenuti fin dai primi dragaggi degli abissi marini, la loro fragilità ha di volta in volta portato a confonderne la reale natura di Protozoi con frammenti di spugne, cnidari, briozoi e di altri organismi ancora. Poiché non è stato finora possibile coltivarli in laboratorio, le conoscenze sulla loro biologia sono praticamente nulle.
Dinozoa
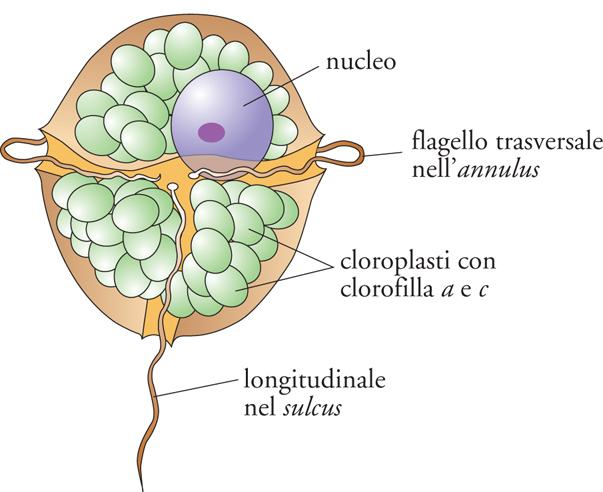
I dinoflagellati sono in larga maggioranza rappresentati da specie planctoniche, sia autotrofe, con un ruolo di primo piano nella produttività primaria del mare, sia eterotrofe, capaci di procurarsi nutrimento (diatomee e altri organismi planctonici) mediante l'uso combinato di un peduncolo protrudibile e di una membrana avviluppante (pallium). Alcune specie, in particolare di Zooxanthella e Symbiodinium, usualmente conosciute come 'zooxantelle', sono comunque divenute endosimbionti mutualisti, essenziali per la vita di molti organismi marini (radiolari, foraminiferi, poriferi, e, soprattutto, coralli ermatipici costruttori delle barriere coralline in cui favoriscono la precipitazione del carbonato di calcio per la costruzione dello scheletro). Le forme libere sono dotate di un'armatura o teca formata da piastre cellulosiche di vario spessore e disegno, che sono sintetizzate all'interno di alveoli (amfiesmi) associati alla faccia interna della membrana plasmatica (e ritenuti omologhi agli alveoli del cortex dei ciliati e apicomplessi, come indica la comune denominazione di 'alveolati' sotto la quale ora si riuniscono questi tre gruppi di protisti). Questa teca, spesso ulteriormente ispessita dall'apposizione di altre ornamentazioni di natura esogena, si presenta tipicamente solcata da due docce: una equatoriale (annulus), contenente un flagello trasversale (e che suddivide la teca stessa tra epiteca e ipoteca), e l'altra subequatoriale (sulcus), contenente un flagello longitudinale (fig. 8).
Al variare di particolari condizioni ambientali, o anche in seguito al deflusso in mare di acque piovane ricche di inquinanti organici, alcune specie di dinoflagellati possono svilupparsi fino a concentrazioni di un milione di cellule/litro e occupare superfici di centinaia di m2. Queste esplosioni demografiche, conosciute anche come 'maree rosse' e 'bioluminescenza del mare' (dovuta in particolare a Noctiluca miliaris), possono avere un forte impatto ambientale per la produzione di tossine estremamente potenti e dannose sia per Crostacei e Bivalvi, che si nutrono dei dinoflagellati che le producono, sia per chi mangia questi organismi inquinati. Nell'uomo, gli effetti neurotossici di tali tossine (denominate 'saxitossine' e 'brevetossine' in relazione a Gymnodinium breve da cui sono state originariamente caratterizzate) sono dovuti a un blocco della pompa Na/K delle cellule nervose e descritti come PSP, Paralytic shellfish poisoning, e NSP, Neurotoxic shellfish poisoning.
Il nucleo dei dinoflagellati (dinokaryon) contiene cromosomi che sono permanentemente in uno stato condensato, privi di nucleoproteine basiche (istoni), e ripartiti alle cellule figlie con l'intervento della membrana nucleare e non dei microtubuli del fuso mitotico. È ora chiaro che queste eccentricità cromosomiche sono di origine secondaria; per lungo tempo, tuttavia, è stato dato loro risalto per sostenere (erroneamente) sia un'organizzazione molto primitiva del nucleo dei dinoflagellati, sia una posizione evolutiva di questi organismi (che sono stati denominati anche 'mesocarioti') intermedia tra procarioti ed eucarioti.
Varie specie sessuate producono zigoti soggetti a perdere rapidamente i loro flagelli e sviluppare una spessa parete cistica, che ne permette la trasformazione in uno stadio di latenza capace di durare anni prima di liberare nuovamente (in seguito al compimento di una meiosi) quattro cellule biflagellate che iniziano una nuova fase moltiplicativa della specie. Queste cisti zigotiche (istricosfere) sono responsabili della ricorrenza delle maree rosse a distanza di anni; inoltre esse rafforzano la capacità di sopravvivenza delle specie tossiche e rappresentano i reperti fossili di specie estinte (ne sono stimate intorno a 5000) risalenti a più di 200 milioni di anni fa (in pratica, al Triassico).
Ciliophora
I ciliati sono cosmopoliti e ubiquitari, e rappresentano una componente costante e spesso preponderante di ogni habitat e catena trofica, sia bentonica sia planctonica. Con le loro quasi 10.000 specie descritte, essi costituiscono il più numeroso raggruppamento di protisti viventi. Oltre alle forme a vita libera largamente preponderanti, ne esistono numerose sia ectocommensali su invertebrati acquatici, sia endocommensali nell'intestino di Vertebrati (come nel caso degli entodiniomorfi, ospiti permanenti dello stomaco di tutti i ruminanti, dove contribuiscono in modo determinante alla digestione della cellulosa). Patogeni sono solo Balantidium e Ichthyophthirius, quest'ultimo responsabile di una malattia spesso mortale (nota come 'muffa bianca') in vari pesci di acqua dolce e d'allevamento. Tra tutti i protisti, i ciliati sono quelli strutturalmente più complessi, soprattutto per quanto riguarda l'organizzazione dell'apparato nucleare e della superficie cellulare. L'apparato nucleare è eterocariote, cioè costituito da nuclei strutturalmente e funzionalmente diversi, noti come 'micronuclei' (o 'nuclei germinali') e 'macronuclei' (o 'nuclei somatici'): i primi sono sferici, diploidi, trascrizionalmente inerti e deputati a generare i nuclei gametici attraverso meiosi in occasione di ogni processo sessuale; i secondi sono polimorfi, poliploidi (eccetto che nelle forme più antiche dei cariorelittidi), attivi nella trascrizione genica, e si differenziano dai prodotti di divisione del synkarion formato dalla fusione dei nuclei gametici.
Nella superficie cellulare, usualmente designata come cortex e spessa da 1 a 4 μm, si distinguono una pellicola e una infraciliatura. La pellicola comprende la membrana plasmatica con annesso un complesso di alveoli, spesso contenenti materiale calcificato per la stabilità meccanica dell'intera superficie cellulare. L'infraciliatura è invece rappresentata dall'insieme dei cinetosomi della cellula e delle strutture che vi sono fisicamente connesse, cioè le ciglia e i fasci di fibrille e microtubuli che ne costituiscono le 'radici'. Questa infraciliatura si organizza nella costruzione di due distinte regioni corporee: una somatica e l'altra orale. La prima è destinata alla locomozione ed è quindi usualmente più estesa e dotata di ciglia singole o associate (come negli ipotrichi) a formare cirri che funzionano come tanti piccoli arti adatti per la locomozione e l'adesione al substrato, più che per il nuoto. La seconda è destinata all'alimentazione e quindi contiene il citostoma usualmente circondato da vari tipi di strutture ciliari composte (membranelle) deputate alla cattura del cibo (batteri, alghe, diatomee). Questa regione orale si differenzia in posizione apicale nelle forme più antiche, ventrale in quelle più moderne, o può andare incontro a regressione completa, come avviene negli acineti che si nutrono per mezzo di tentacoli succhiatori.
Anche in relazione alla sessualità, i ciliati appaiono piuttosto eccentrici rispetto a tutti gli altri protisti, per il fenomeno della coniugazione che hanno sviluppato. Questo fenomeno, infatti, comporta la formazione di coppie tra due cellule che compiono un mutuo scambio di nuclei gametici (derivati per meiosi del loro micronucleo) e formano ciascuna un synkaryon (o nucleo zigotico, destinato a ricostituire un nuovo apparato nucleare) per poi separarsi e riprendere la loro vita trofica vegetativa. Nelle forme più evolute, la coniugazione si manifesta sotto il controllo di sistemi di mating type controllati geneticamente e distinti tra binari e multipli, a seconda che il numero di mating type che li compone sia due, o più di due. Per mating type si intendono popolazioni clonali di cellule (cioè popolazioni ciascuna originata per moltiplicazione asessuale da un unico capostipite) che sono, allo stesso tempo, capaci di formare coppie di coniuganti in seguito a mutue interazioni mediate da contatto fisico, o da segnali chimici diffusi nell'ambiente (noti come 'feromoni'), e refrattarie a formarne al loro interno.
Apicomplexa
Tutte le specie di questo raggruppamento sono parassite, prive di qualsiasi organulo locomotorio (solo lo stadio di microgamete può sviluppare uno o due flagelli) e uniformemente delimitate da un complesso di tre membrane, delle quali le due sottostanti la membrana plasmatica sono interpretate come membrane alveolari (cioè equivalenti a quelle che delimitano gli alveoli dei ciliati e gli amfiesmi dei dinoflagellati) e si interrompono solo localmente per permettere le attività eso- e endocitotiche della cellula.
La denominazione di 'apicomplessi' deriva da una struttura composta da più subunità concentrate nella regione anteriore degli stadi infettivi (sporozoiti e merozoiti), che è nota appunto come 'complesso apicale' ed è visibile solo a livello ultrastrutturale. Nelle forme intracellulari (coccidi, emosporidi e piroplasmi), questo complesso permette la penetrazione dello stadio infettivo nelle cellule ospiti, mentre in quelle extracellulari (cioè le gregarine) ne consente l'adesione alle pareti cellulari. Tra le subunità che ne fanno parte, un ruolo funzionale centrale è giocato da un complesso di 2-5 organuli a forma di clava (roptrie), che secernono una batteria di enzimi ad azione litica e cooperano con l'attività secretoria di numerosi granuli citoplasmatici nella costruzione di un vacuolo parassitoforo, che protegge con una sua propria membrana il parassita dall'attività lisosomale (difensiva) della cellula ospite. Oltre al complesso apicale, gli apicomplessi possiedono un solo nucleo vescicolare, un unico grosso mitocondrio a forma di Y e un organulo di origine plastidiale (apiplasto), verosimilmente originato da un'alga rossa endosimbionte, che possiede un suo proprio DNA circolare coinvolto nel controllo della biosintesi degli acidi grassi della cellula.
Gli apicomplessi presentano tutti cicli biologici tipicamente articolati in un'alternanza di tre distinte fasi, due delle quali asessuali (sporogonia e schizogonia) e l'altra sessuale (gamogonia). La fase sporogonica si compie a carico degli zigoti, dalla cui divisione si formano gli sporozoiti, che nelle gregarine e nei coccidi penetrano nell'intestino dell'ospite protetti in oocisti e/o sporocisti, mentre negli emosporidi e piroplasmi sono direttamente inoculati da un artropode vettore all'interno dei vasi sanguigni. Nell'ospite, gli sporozoiti si nutrono come trofozoiti e danno luogo alla fase schizogonica moltiplicandosi in merozoiti (che sono stati spesso descritti in specie di interesse medico e veterinario come zoiti, schizozoiti, bradizoiti, tachizoiti, dormozoiti, cistozoiti e così via, in relazione a varie peculiarità morfologiche e/o comportamentali). Questi ultimi invadono nuove cellule dell'ospite e ripetono altre fasi schizogoniche finché, usualmente in coincidenza con il passaggio a un secondo ospite, alcuni si differenziano sessualmente e compiono la fase gamogonica, dividendosi per produrre gameti maschili più piccoli e flagellati (microgameti) e gameti femminili grossi e immobili (macrogameti), dalla cui fecondazione avranno origine gli zigoti che iniziano un nuovo ciclo.
A livello evolutivo, gli apicomplessi includono le gregarine che ne costituiscono le forme più primitive (ca. 1500 specie), tutte viventi in cavità corporee (nel lume intestinale in particolare) di invertebrati. I loro trofozoiti presentano grosse dimensioni (che possono raggiungere anche 3÷4 mm), sono chiaramente distinti da un setto trasversale in una regione anteriore (protomerite) e in una regione posteriore contenente il nucleo (deutomerite); essi sono deputati a produrre gameti in seguito a una loro particolare unione a tandem (sizigia).
Evolutivamente più recenti e con sporozoiti che invadono le cellule epiteliali e del sangue dei Vertebrati sono invece i coccidi (i più rappresentati, costituiti da circa 2000 specie), gli emosporidi e i piroplasmi. Tra i coccidi ritroviamo forme estremamente patogene sia per gli animali da allevamento (soprattutto uccelli), sia per l'uomo. Le prime sono rappresentate da specie di Eimeria e Isospora, causa di malattie letali comunemente note come 'coccidiosi'; le seconde da Cryptosporidium parvum (responsabile di gravi dissenterie), che contamina i bacini d'acqua dolce con le sue oocisti molto resistenti espulse con le feci da vari Mammiferi suoi ospiti, e Toxoplasma gondii, il cui ospite definitivo è il gatto (oppure un altro felino), mentre l'uomo rientra nella vasta gamma di ospiti intermedi. Oltre che ingerendo oocisti mediante contatto con il gatto, l'uomo può infettarsi di T. gondii anche attraverso l'ingestione di carne contenente pseudocisti. Fortunatamente per l'uomo, la risposta anticorpale è generalmente forte per cui, salvo che negli individui immunodepressi o immunodeficienti, i danni provocati da questo parassita restano in genere limitati a un'infiammazione dei linfonodi. È invece la trasmissione di cisti al feto per via transplacentare che genera serie patologie; particolarmente quando questa trasmissione si verifica in occasione della prima gravidanza si hanno forti incidenze sia di aborto sia di gravi danni cerebrali (idrocefaliti) nei nati ai primi mesi di vita.
Gli emosporidi vivono tutti a spese dei globuli rossi di Rettili, Uccelli e Mammiferi (soprattutto Roditori e Primati) giacché vi compiono la fase schizogonica del ciclo. La loro trasmissione è usualmente opera di una zanzara del genere Anopheles, Aedes o Culex, nella quale essi invece completano sia la fase sessuale sia la sporogonia. In relazione a questo tipo di trasmissione diretto, gli sporozoiti degli emosporidi non sono mai protetti in oocisti o sporocisti. Di tutti gli emosporidi, il più conosciuto è Plasmodium, responsabile nell'uomo della malaria; la sua specie più patogena, P. falciparum, causa non meno di due milioni di morti ogni anno e interessa il territorio di oltre cento nazioni. Gli sporozoiti di Plasmodium passano dal lume delle ghiandole salivari di un esemplare infetto di Anopheles direttamente nel sangue dell'uomo con la puntura. Gli sporozoiti prima si moltiplicano nelle cellule epatiche; ogni merozoite prodotto da questa fase eso-eritrocitica penetra poi in un eritrocita, digerendone l'emoglobina e determinandone la rottura per immettere di nuovo nel circolo sanguigno i prodotti della sua moltiplicazione schizogonica, in media una quindicina di altri merozoiti, destinati ad aggredire altri eritrociti. La rottura pressoché sincrona degli eritrociti infetti determina un accumulo di cataboliti tossici nel sangue, cui si devono gli eccessi febbrili ricorrenti tipici della malaria (ogni due o tre giorni, a seconda che si tratti della 'terzana' o 'quartana' e considerando il giorno dell'eccesso come primo giorno). Dopo che si sono compiute almeno quattro o cinque fasi schizogoniche eritrocitarie, una parte dei merozoiti si differenzia sessualmente per generare i macro- e microgameti. Questo differenziamento tuttavia si compie solo in seguito al trasferimento di questi merozoiti dal sangue dell'uomo nell'intestino della zanzara. È infatti la diminuzione di temperatura corporea che c'è tra l'uomo e la zanzara che lo induce. I macrogameti fecondati divengono zigoti mobili (oocineti), capaci di passare tra una cellula epiteliale e l'altra della parete intestinale della zanzara, per annidarsi immediatamente sotto la membrana esterna, dove compiono la loro fase sporogonica ingrossandosi in oocisti (note anche come 'vermicoli'). In seguito alla rottura della parete, ogni oocisti libera centinaia di sporozoiti, che subito migrano dall'emocele della zanzara nel lume delle ghiandole salivari dove sostano in attesa di essere inoculati in un nuovo ospite. I piroplasmi si identificano essenzialmente con alcune decine di specie di Babesia e Theileria, responsabili di gravi patologie soprattutto nei bovini, in cui si compie la fase schizogonica a carico sia degli eritrociti, sia dei linfociti. La trasmissione degli sporozoiti avviene per mezzo della puntura di varie specie di zecche, non solo da parte di individui adulti, ma anche degli stadi ninfali che li ricevono dalla madre per via transovariale e contribuiscono così a espandere enormemente l'infezione.
Microsporidia
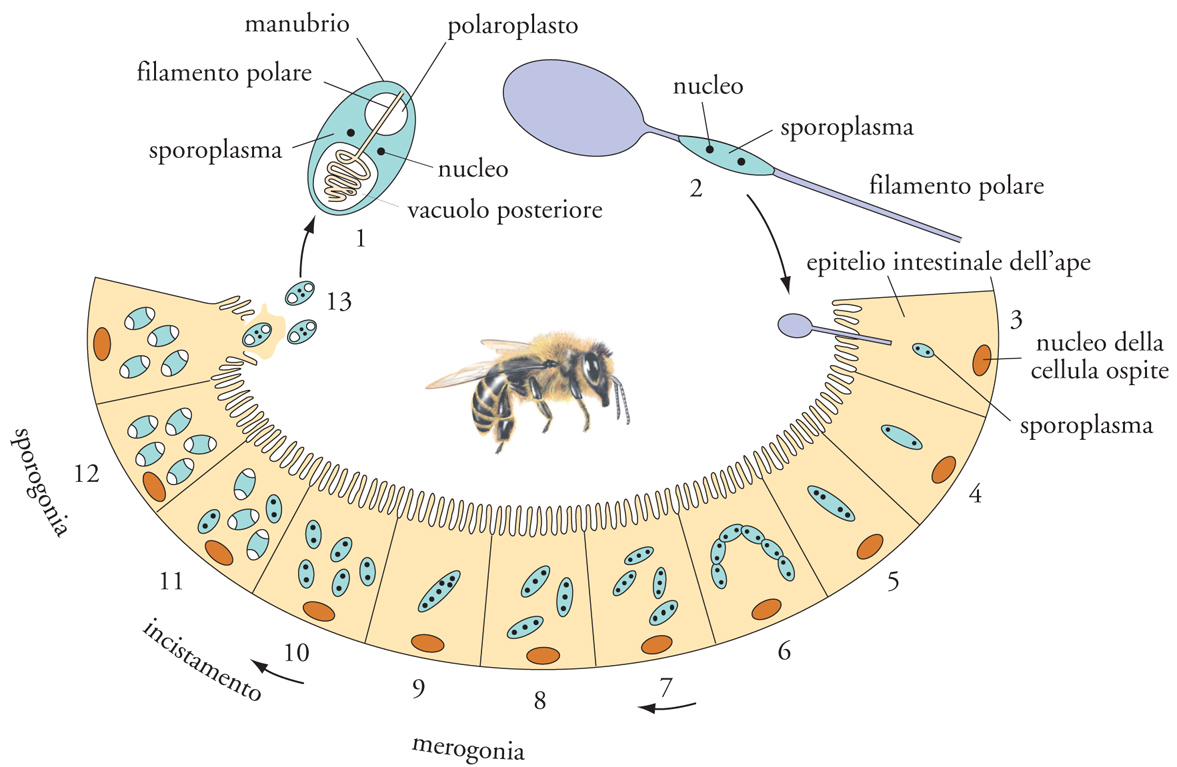
I microsporidi sono parassiti obbligatori intracellulari di molti invertebrati e di tutti i Vertebrati, a ogni modo con larga predilezione per Artropodi (fig. 10) e Pesci. L'assenza di mitocondri e un'organizzazione dei ribosomi di tipo più procariotico che eucariotico li hanno per vario tempo fatti ritenere molto primitivi, in pratica uno dei primi gruppi nell'evoluzione dei protisti. Analisi genetiche molecolari hanno poi definitivamente disconosciuto questa ipotesi, chiarendo l'origine secondaria di queste peculiarità e la sostanziale natura fungina dei microsporidi. Le loro spore sono spesso binucleate e dotate di una doppia parete, proteica esterna e chitinosa interna, a protezione di un citoplasma che costituisce il reale agente infettivo (sporoplasma), la cui estrusione nella cellula ospite coinvolge un complesso di distinte strutture, ognuna delle quali con una sua specifica funzione: un cappuccio polare per l'ancoraggio alla superficie della cellula ospite; un filamento polare, spiralato e connesso con un manubrio al cappuccio polare, per scaricare lo sporoplasma all'inerno della cellula ospite; un polaroplasto formato da un complesso di membrane fenestrate che si rigonfia, di concerto con il rigonfiarsi anche di un vacuolo posteriore, per generare pressione sul filamento e facilitarne lo svolgimento.
Nella cellula ospite, lo sporoplasma si accresce rapidamente e si moltiplica in sporoblasti destinati a differenziarsi in nuove spore, le più precoci delle quali germinano rapidamente nell'arco di 1-2 giorni per propagare l'infezione dentro lo stesso ospite, mentre quelle più tardive germinano in tempi più lunghi e vanno a colonizzarne di nuovi. Questo tipo di ciclo molto semplificato si applica alle specie definite 'monomorfiche' (od 'omosporiche'), come Nosema bombycis (parassita del baco da seta), che producono spore di un solo tipo, specifiche per un solo ospite. Altre specie, invece, definite 'polimorfiche' (o 'eterosporiche'), come quelle di Amblyospora, ne producono diversi tipi capaci di infettare più di un ospite.
Myxozoa
I mixozoi sono tutti endoparassiti obbligatori essenzialmente di Pesci, tra i Vertebrati, e di platelminti, e anellidi, tra gli invertebrati. La loro inclusione qui tra i protisti segue solo ragioni pratiche legate alla consuetudine di comprendere i mixozoi, insieme ai microsporidi e gli attuali apicomplessi, sotto la comune denominazione di sporozoi. Analisi molecolari tuttavia indicano chiaramente che le loro parentele evolutive sono con gli cnidari, con le narcomeduse in particolare. L'organizzazione inequivocabilmente pluricellulare delle loro spore è inoltre nota fin dall'inizio del Novecento. Ognuna è infatti formata da un complesso di sporoplasmi infettivi, valve e capsule polari, che sono del tutto simili alle nematocisti degli cnidari e dotate di filamento polare per scaricarsi. La conoscenza completa del ciclo vitale dei mixozoi è relativamente recente e si riferisce essenzialmente al ciclo di Myxobolus cerebralis che si svolge tra trote e oligocheti del genere Tubifex, e si è quindi prestato (per la facile reperibilità di questi due ospiti) a essere più facilmente analizzato sperimentalmente anche in laboratorio. Le trote si infettano con le spore di Myxobolus ingerendole con un verme che le contiene, o venendoci a contatto a livello delle branchie se queste spore si disperdono nell'ambiente in seguito a morte di vermi infetti. Gli sporoplasmi ameboidi, liberati da questo primo tipo di spore (actinospore), migrano nelle cellule cartilaginee e del sistema nervoso centrale della trota, dove producono cellule generative capaci di invadere nuove cellule prima di differenziare uno sporoblasto. Da questo si sviluppano tre diversi tipi di cellule (valvogeniche, capsulogeniche e sporoplasmiche), che si associano a formare un secondo tipo di spore (mixospore), destinate a essere espulse dalla trota con le feci o in seguito a morte. Se ingerite da un nuovo verme, queste spore liberano a loro volta sporoplasmi che, negli spazi intercellulari dell'epitelio intestinale, finiscono per generare un complesso di due cellule germinali e due cellule somatiche. Le prime compiono la meiosi e producono gameti maschili e femminili; le seconde si dividono in otto cellule destinate a proteggere gli zigoti che si originano dalla fecondazione dei gameti e generano sporoblasti, da cui si riformano le actinospore che si liberano dal verme per propagare l'infezione ad altre trote. Alla luce di queste conoscenze sul ciclo di Myxobolus cerebralis, si ritiene che la tradizionale suddivisione dei Myxozoa tra specie (asessuate) con ciclo nei Pesci (Myxosporea) e specie (sessuate) con ciclo negli oligocheti acquatici (Actinosporea) sia sostanzialmente errata, e che sia invece corretto generalizzare dicendo che, complessivamente, i mixozoi richiedono sia Pesci sia oligocheti per completare il loro ciclo biologico.
Bibliografia
Anderson 1983: Anderson, Orvil R., Radiolaria, Berlin, Springer, 1983.
Anderson 1987: Anderson, Orvil R., Comparative protozoology. Ecology, physiology, life history, Berlin, Springer, 1987.
Corliss 1979: Corliss, John O., The ciliated protozoa: characterization, classification and guide to the literature, Oxford, Pergamon, 1979.
Gall 1986: Gall, Joseph G., The molecular biology of ciliated protozoa, Orlando, Academic Press, 1986.
Grell 1973: Grell, Karl G., Protozoology, 3. ed., Berlin-Heidelberg-New York, Springer, 1973.
Gupta 2002: Gupta, Barun K.S., Modern Foraminifera, Dordrecht, Kluwer Academic, 2002.
Harrison, Corliss 1991: Harrison, Frederick W. - Corliss, John O., Protozoa, in: Microscopic anatomy of Invertebrates, edited by Frederick W. Harrison, New York, Wiley-Liss, I, 1991.
Hausmann 2003: Hausmann, Klaus - Hülsmann, Norbert - Radek, Renate, Protistology, 3. ed., Berlin, Schweizerbart, 2003.
Lee 2000: The illustrated guide to the Protozoa, 2. ed., edited by John J. Lee e altri, Lawrence (Kan.), Allen, 2000.
Lee, Anderson 1991: Lee, John J. - Anderson, Orvil R., Biology of Foraminifera, London, Academic Press, 1991.
Levine 1988: Levine, Norman D., Protozoan Phylum Apicomplexa, Boca Raton (Fla.), CRC, 1988.
Margulis 1990: Handbook of protoctista, edited by Lynn Margulis e altri, Boston, Jones and Barlett, 1990.
Patterson, Larsen 1991: The biology of free-living heterotrophic flagellates, edited by David J. Patterson, Jacob Larsen, Oxford, Clarendon, 1991.
Sleigh 1989: Sleigh, Michael A., Protozoa and other protists, 2. ed., London, Arnold, 1989.
© Istituto della Enciclopedia Italiana - Riproduzione riservata


