Radiazione
Enciclopedia on line
Energia che si propaga per onde (r. ondulatoria) o per corpuscoli (r. corpuscolare) e anche il fenomeno stesso dell’emissione, dell’irradiamento di tale energia (r. di una sorgente, r. di un’antenna, diagramma di r.).
Fisica
Tipiche r. ondulatorie sono le r. sonore, tipiche r. corpuscolari le r. alfa, beta ecc. La r. elettromagnetica (in particolare la r. luminosa), che può essere descritta come la propagazione sia di onde elettromagnetiche, sia di fotoni (corpuscoli con massa nulla), è classificabile come r. ondulatoria o come r. corpuscolare, a seconda della rappresentazione più adeguata al fenomeno allo studio.
Il termine r. trae origine dalla circostanza che la propagazione dell’energia può pensarsi svolgentesi lungo raggi o costituita da raggi, essendo questi ultimi le direzioni di propagazione delle onde o dei corpuscoli anzidetti. Spesso la specificazione si riferisce a determinate proprietà o all’origine della r.: r. penetranti, r. radioattive (➔ radioattività), r. cosmica, r. di fondo cosmico, r. di frenamento, r. atmosferiche.
R. ionizzanti e non ionizzanti
Le r. ionizzanti sono quelle che producono la ionizzazione direttamente per collisione con gli elettroni del mezzo, come le particelle cariche veloci (elettroni, protoni ecc.), o per effetto fotoelettrico ed effetto Compton nel caso dei fotoni sufficientemente energetici (convenzionalmente con energia maggiore di 10 eV e quindi con lunghezza d’onda minore di 10–7 m), oppure indirettamente attraverso la produzione di particelle cariche come nel caso dei neutroni (che danno luogo a reazioni nucleari) o dei fotoni con energia maggiore di 1 MeV (che danno luogo alla creazione di coppie). Le r. non ionizzanti sono quelle che non producono la ionizzazione, come le r. sonore o quelle elettromagnetiche con lunghezza d’onda maggiore di 10–7 m.
R. cosmica
R., costituita da particelle elementari e nuclei atomici (detti anche raggi cosmici), che raggiunge la Terra, provenendo dallo spazio. Essa ha origine in parte nella Via Lattea (r. cosmica galattica) e in parte al di fuori di questa (r. cosmica extragalattica). Una frazione della componente galattica è generata nel Sistema solare: tuttavia, queste particelle di origine ‘locale’, irradiate soprattutto dal Sole (raggi cosmici solari), vengono di solito considerate separatamente (➔ Sole).
Raggi cosmici primari e secondari
Le energie dei raggi cosmici coprono un ampio intervallo, che si estende da ∼107 eV fino a ∼1020 eV, un valore che supera largamente quello di qualsiasi altra r. nota. A causa della loro elevatissima energia, le particelle che entrano nell’atmosfera, i raggi cosmici primari, interagiscono con i nuclei atomici dei gas atmosferici generando una varietà di altre particelle e fotoni, che costituiscono i raggi cosmici secondari. Lo studio di queste interazioni ha avuto, nel passato, una grande rilevanza per la fisica delle alte energie: infatti, molte particelle elementari (per esempio, il positrone e i mesoni π, μ e K) sono state scoperte proprio investigando la r. cosmica. Con lo sviluppo degli acceleratori di particelle, che consentono di effettuare esperimenti in condizioni controllate, lo studio dei raggi cosmici ha perduto interesse da questo punto di vista, ma ha acquistato una crescente importanza astrofisica e cosmologica: basti pensare che i raggi cosmici rappresentano gli unici campioni di materia che giungano a noi dall’esterno del Sistema solare.
Proprietà osservative della r. cosmica primaria
La natura dei raggi cosmici primari cominciò a essere investigata in modo diretto negli anni 1940 con esperimenti su pallone che portavano i rivelatori (contatori Geiger e, in seguito, emulsioni nucleari) a quote di oltre 20.000 metri. Le osservazioni spaziali hanno avuto inizio negli anni 1960 con i satelliti americani delle serie IMP e OGO e i sovietici Proton. Questi esperimenti hanno dato un contributo importantissimo alla conoscenza della r. cosmica perché hanno reso possibile effettuare misure di lunga durata e non contaminate dall’atmosfera.
Si è stabilito che la r. primaria è costituita per ∼90% da protoni. La parte residua consiste di nuclei di elementi più pesanti dell’idrogeno e di una piccola frazione (∼1%) di elettroni. La composizione chimica della r. cosmica è all’incirca costante nell’intervallo di energie da ∼200 MeV a 1-2 GeV. D’altro canto, per la maggior parte degli elementi non vi sono grosse discrepanze fra le abbondanze dei raggi cosmici e quelle del Sistema solare. Tuttavia, nei raggi cosmici si osserva la tendenza a ‘riempire’ i buchi delle abbondanze di certi elementi che scarseggiano nel Sistema solare: i casi più vistosi sono quelli dei metalli leggeri (litio, berillio e boro) e degli elementi (dallo scandio al manganese) che precedono il ferro. Il fenomeno è spiegato dalle collisioni fra particelle che avvengono nello spazio interstellare, nelle quali i nuclei massicci si frammentano, creando nuclei più leggeri (le cosiddette reazioni di spallazione). Oltre il ferro (il nucleo più stabile), che presenta un picco marcato, le abbondanze degli elementi decrescono rapidamente sicché non si riesce a determinarle con precisione: tuttavia, qualitativamente, si è accertata la presenza nella r. cosmica di quasi tutti gli elementi più pesanti fino all’uranio e si è riusciti a risolvere la distribuzione delle abbondanze isotopiche di parecchi elementi.
Altre osservazioni importanti sono quelle delle antiparticelle. Si è trovato che i positroni sono ∼10 volte meno numerosi degli elettroni. Ciò indica che gli elettroni osservati non provengono, per la maggior parte, da collisioni di nuclei nello spazio interstellare: in questo caso, infatti, essi dovrebbero essere altrettanto numerosi quanto i positroni, dal momento che le due particelle sarebbero prodotte in coppie nel decadimento dei mesoni, attraverso la catena π<μ<e±. Nella r. cosmica sono stati rivelati anche antiprotoni, ma non è ancora chiaro se la loro abbondanza sia o no maggiore di quella prevista in base alla frequenza delle collisioni nucleari in cui essi sono generati. Non sono invece stati individuati antinuclei di elementi più pesanti dell’idrogeno: sarebbe di grande interesse scoprirli perché ciò indicherebbe l’esistenza, nell’Universo, di estese regioni di antimateria.
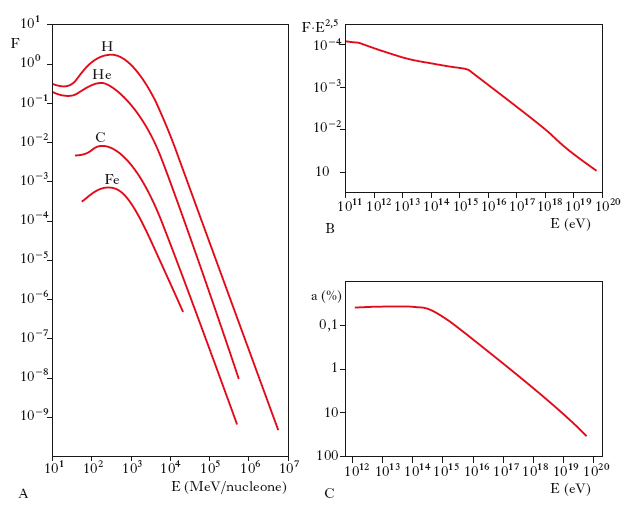
La fig. 1A mostra gli spettri di energia di alcuni dei nuclei più abbondanti nella r. cosmica. Per confrontare gli andamenti delle diverse specie, la scala delle ascisse è espressa in MeV/nucleone (cioè l’energia è divisa per il numero di massa del nucleo). La flessione delle curve alle basse energie è un fenomeno locale, dovuto al fatto che le particelle meno energetiche non riescono a penetrare nella eliosfera. Alle alte energie, tutti gli spettri hanno pressoché lo stesso andamento, descritto da una legge di potenza del tipo E–γ, con γ=2,6-2,7. Il valore γ è, tuttavia, sensibilmente diverso per le specie nucleari (che non appaiono in figura), generate attraverso le reazioni di spallazione. Lo spettro di energia complessivo dei raggi cosmici è stato misurato su un intervallo assai più esteso di quello che è possibile ottenere per ciascuna specie nucleare separatamente. Si trova che la pendenza dello spettro cambia intorno a 1016 eV, divenendo più ripida a energie maggiori (fig. 1B). Per E>1019 eV, lo spettro tende nuovamente ad appiattirsi. Lo spettro di energia degli elettroni si estende fino a ∼1012 eV ed è descritto da una legge di potenza con γ=2,6. La densità di energia dei raggi cosmici, nelle vicinanze del Sistema solare, risulta essere ∼1 eV/cm3. Se ne deduce che il flusso di energia, trasportato da questa r., che incide sulla Terra, è dello stesso ordine di grandezza di quello associato alla luce delle stelle.
La r. cosmica è quasi perfettamente isotropa, cioè il suo flusso è all’incirca lo stesso in tutte le direzioni. Per E<1014 eV, l’anisotropia (fig. 1C) è dell’ordine dello 0,05%. A energie maggiori essa aumenta gradualmente, raggiungendo un valore di ∼10% per E∼1019 eV. Solo per E>1019 eV il flusso diventa fortemente collimato.
Origine della r. cosmica
Il problema dell’origine della r. cosmica non è stato ancora completamente risolto. Vi sono, comunque, buone ragioni per ritenere che la maggior parte delle particelle con E<1019 eV sia generata nella Via Lattea. Origine galattica hanno certamente gli elettroni. Valutando le perdite di energia subite dagli elettroni più energetici (E∼1012 eV) per effetto Compton nelle collisioni con i fotoni della r. di fondo cosmico, si trova che essi percorrono nel mezzo interstellare un cammino l≲0,3 Mpc. D’altra parte, la quasi isotropia del flusso implica che le loro traiettorie siano tortuose. Più precisamente, è: δ=r/l, dove δ è il valore dell’anisotropia, l il cammino percorso e r la corrispondente distanza in linea retta. Per δ=0,5∙10–3 e l=0,3 Mpc, si ha r=0,1 kpc, che è una distanza assai più piccola delle dimensioni del disco galattico. Se ne deduce che gli elettroni devono provenire da sorgenti relativamente vicine, situate nel disco galattico. Per quanto riguarda i protoni e i nuclei più pesanti, la loro origine galattica è comprovata con argomenti meno diretti. Se le particelle sono generate nella Via Lattea e confinate in essa dal campo magnetico interstellare, la loro densità di energia non potrà superare quella del campo. In accordo con questa aspettativa, si trova che entrambe le densità di energia sono dello stesso ordine di grandezza (∼1eV/cm3). All’aumentare dell’energia delle particelle, si attendono due effetti. Da una parte, esse sfuggiranno più facilmente e quindi il loro tempo di confinamento nella Via Lattea sarà più breve; dall’altra, le irregolarità del campo magnetico, che diffondono le particelle tendendo a rendere isotropa la loro distribuzione, diventeranno meno efficaci sicché l’anisotropia aumenterà. Entrambi i fenomeni sono osservati: per energie maggiori di 1015-1016 eV, il flusso delle particelle subisce una brusca diminuzione, come indicato dall’aumento di pendenza dello spettro (fig. 1B), mentre l’anisotropia cresce (fig. 1C).
Un’analisi quantitativa conferma che sia l’andamento dello spettro energetico che quello dell’anisotropia sono spiegati in modo soddisfacente, fino a energie di ∼1019 eV, assumendo che la r. cosmica sia confinata nella nostra Galassia. Vi sono, d’altra parte, altre osservazioni, concernenti la composizione chimica e isotopica della r., che indicano che la regione di confinamento deve essere più vasta del disco galattico. L’abbondanza dei metalli leggeri (Li, Be, B), prodotti nelle reazioni di spallazione, implica, infatti, che i raggi cosmici abbiano attraversato uno spessore di materia pari a ∼5 g/cm2. Inoltre, poiché l’abbondanza dell’isotopo radioattivo 10Be rispetto al berillio ordinario è minima, essi devono aver viaggiato per tempi almeno 10 volte più lunghi del periodo di dimezzamento di questo isotopo (2∙106 anni). Combinando le due osservazioni precedenti, si deduce che la densità media del mezzo attraversato dalle particelle è minore di 0,5∙10–24 g/cm3, un valore significativamente più basso della densità media del gas nel disco galattico (∼2∙10–24 g/cm3). Si conclude che la r. deve essere confinata a una regione di spazio avente un volume parecchie volte maggiore del disco galattico. Presumibilmente, dunque, i raggi cosmici permeano non soltanto il disco, ma anche l’alone della Via Lattea. Nella Galassia, vi sono svariate sorgenti di particelle energetiche, che possono alimentare la r. cosmica. Sembra però che il contributo maggiore provenga dalle supernove e dai loro residui (le pulsar). Da un punto di vista energetico, le supernovae sono senz’altro sorgenti adeguate. Tenendo conto delle dimensioni della Galassia e dei tempi di confinamento delle particelle, si trova che per alimentare la r. cosmica occorre un’energia di ∼1041 J/anno. Dal momento che nella Via Lattea avvengono mediamente 2 esplosioni di supernove ogni secolo e che in ciascuna di esse si libera un’energia di ∼1044 J, sarebbe sufficiente che il 10% di questa energia andasse ad accelerare le particelle per giustificare i flussi osservati.
Si pensa che le particelle più energetiche della r. cosmica (E>1019 eV) non siano generate nella Via Lattea. A tali energie, il raggio di girazione di un protone nel campo magnetico interstellare è dello stesso ordine delle dimensioni della Galassia. Pertanto, la particella percorre una traiettoria quasi rettilinea e la sua direzione di provenienza può darci un’indicazione della sorgente che l’ha prodotta. Si trova che il flusso dei raggi cosmici, a queste energie estreme, è fortemente anisotropo (fig. 1C) e proviene, entro ∼10°, dalla direzione in cui si trova l’ammasso di galassie della Vergine (cioè non lontano dal polo nord galattico). D’altra parte, dal punto di vista energetico, è del tutto plausibile che le particelle siano generate in questo ammasso: infatti, per spiegare il flusso osservato, è sufficiente supporre che ciascuna galassia dell’ammasso contribuisca all’alimentazione della r. con un’energia di 1040 J/anno, un valore che è 10 volte più piccolo dell’energia necessaria a sostenere il flusso di raggi cosmici nella Via Lattea.
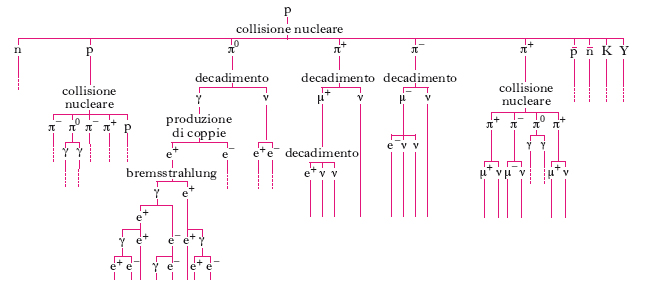
La r. cosmica secondaria
Quando entrano nell’atmosfera, i raggi cosmici primari danno luogo a catene di interazioni nucleari, il cui risultato è la formazione della r. cosmica secondaria. In media, i protoni subiscono la prima collisione con un nucleo dei gas atmosferici dopo avere attraversato uno spessore di ∼70 g/cm2 di materia (cioè 1/15 della massa totale di aria al di sopra del livello del mare); le particelle α (elio) e i nuclei più pesanti collidono ancora prima. Di conseguenza, la probabilità che una particella dei raggi cosmici primari sfugga a collisioni nucleari finché non raggiunge il livello del mare è praticamente nulla. Quando una particella primaria (per fissare le idee, un protone) urta contro un nucleo, in generale lo disintegra, dando inizio ai processi descritti schematicamente in fig. 2. Fra i prodotti della collisione, vi sono protoni e neutroni (che possono avere energie sufficienti per rompere, a loro volta, altri nuclei) e particelle di vita breve, come mesoni π, mesoni K e iperoni. I più numerosi sono i mesoni π. La storia successiva di queste particelle dipende dalla loro carica. I mesoni π0 (neutri) hanno una vita media brevissima (2∙10–16 s) sicché, per la massima parte, decadono in fotoni senza partecipare ad altre collisioni. I fotoni danno luogo a un processo a catena: essi si materializzano in coppie di elettroni e positroni, che, a loro volta, irraggiano nuovi fotoni. Si sviluppa così una cascata elettrofotonica. Man mano che questa si ingrossa, l’energia media dei fotoni e delle particelle (elettroni e positroni) che la costituiscono diminuisce, sicché, a un certo punto, diventa insufficiente a proseguire il processo. La cascata, allora, si esaurisce. I mesoni π±, avendo una vita più lunga dei π0, hanno discrete probabilità di subire collisioni nucleari prima di decadere, generando un mesone μ± e un neutrino. I mesoni μ hanno una vita media relativamente lunga (2,2∙10–6 s) e un forte potere penetrante, in quanto perdono energia quasi soltanto per ionizzazione. Nel loro insieme, le particelle e i fotoni, discendenti da una medesima particella primaria, costituiscono uno sciame atmosferico.

Sia la composizione che l’intensità complessiva della r. cosmica (primaria e secondaria) variano fortemente con l’altezza. La fig. 3 dà l’andamento del flusso Φ (numero di particelle per cm2 per angolo solido unitario per secondo) con la profondità atmosferica p (misurata in g/cm2 dal culmine dell’atmosfera), per tre diverse componenti della r.: le particelle nuclearmente attive (protoni, neutroni ecc.) con energie maggiori di 1 GeV (curva a); gli elettroni con energie maggiori di 100 MeV (curva b); i mesoni μ con energie maggiori di 200 MeV (curva c). Si nota che, mentre l’intensità della prima componente diminuisce in modo monotono con la profondità, quelle delle altre due dapprima aumentano, raggiungendo un massimo fra 100 e 200 g/cm2, e solo in seguito cominciano a diminuire. D’altra parte, il flusso dei mesoni μ diminuisce più lentamente di quello degli elettroni, sicché al livello del mare esso diventa dominante. Di conseguenza, nella r. cosmica secondaria, si usa distinguere una componente dura, fortemente penetrante, formata dai mesoni μ, da una componente molle, assai meno penetrante, formata dalle cascate elettrofotoniche.
R. gamma cosmica
Nell’ultimo decennio del 20° sec. si è avuto un notevole progresso nello studio del fenomeno legato alla r. gamma cosmica, conosciuto come burst di raggi gamma (gamma ray burst). Questo fenomeno consiste in rapidi, intensi lampi di raggi gamma che illuminano per qualche frazione di secondo il cielo visto in r. gamma. Accadono con una frequenza di circa uno al giorno e quando si manifestano coprono completamente la r. gamma proveniente dalle altre sorgenti usuali, Sole compreso (➔ gammaastronomia).
R. di fondo cosmico
R., costituita da fotoni di corpo nero a temperatura di 2,73 K, che permea l’Universo in modo omogeneo. Secondo la teoria del big-bang (➔), tali fotoni sono stati prodotti nelle primissime fasi evolutive dell’Universo, durante le annichilazioni materia-antimateria: si tratta quindi della r. più antica che oggi si può osservare nel nostro Universo. L’esistenza di questa r. fossile è, insieme all’espansione dell’Universo e al rapporto di densità tra gli atomi di idrogeno e quelli di elio, un punto di forza della teoria del big-bang, che prevede l’origine dell’Universo da uno stato iniziale con temperatura e densità elevatissime; durante la successiva espansione la temperatura e la densità sono diminuite fino a raggiungere i valori attuali. Schematicamente, si può dividere la storia dell’Universo in due periodi separati da un’epoca chiamata era del disaccoppiamento.
Nel primo periodo, la r. e la materia sono fortemente accoppiate e tali da costituire un unico fluido, il plasma primordiale, che si può considerare in equilibrio termodinamico. Infatti, il cammino libero medio dei fotoni nel plasma è molto breve, a causa della diffusione da parte degli elettroni liberi ivi presenti: ne consegue una veloce termalizzazione. In tutta questa prima fase l’Universo è opaco alla r., a causa dell’elevatissimo numero di diffusioni. Al disaccoppiamento, la temperatura è scesa a circa 3000 K, e quindi si formano atomi di idrogeno neutro, si riduce drasticamente il numero di elettroni liberi e l’equilibrio termodinamico non viene più mantenuto. Da questo momento in poi r. e materia evolvono indipendentemente e l’Universo diventa trasparente.
La regione in cui avviene l’ultima diffusione da parte degli elettroni liberi viene detta superficie di ultimo scattering; a causa della sua temperatura la r. presenta la distribuzione spettrale di un corpo nero a 3000 K. In seguito alla successiva espansione dell’Universo la temperatura dello spettro di corpo nero è diminuita in ragione inversa dell’espansione e attualmente è intorno a 2,7 K. Il massimo d’intensità si ha quindi alle lunghezze d’onda millimetriche ed è per questo che si parla di fondo cosmico a microonde. Utilizzando come scala dei tempi il redshift, la superficie di ultimo scattering si trova a redshift z pari a 1000. La r. di fondo nasce proprio da z=1000 e quindi il suo studio permette di avere informazioni sulle condizioni dell’Universo in un’epoca molto remota. Per confronto, si noti che le sorgenti discrete più lontane osservabili oggi sono i quasar rivelati fino a z∼5. L’epoca tra z=5 e z=1000 è di grande interesse cosmologico, perché in questo periodo si sono formate le prime strutture nell’Universo. La r. emessa da questi processi dovrebbe produrre un fondo cosmico infrarosso, non ancora osservato. Il fondo cosmico a microonde si dimostra un ottimo banco di prova per verificare sperimentalmente le teorie cosmologiche. In particolare si studiano il suo spettro e le sue variazioni in intensità in funzione della direzione di osservazione, cioè le sue anisotropie. Una misura accurata dello spettro permette di stabilire se si sono verificate iniezioni di energia e quindi deviazioni dall’equilibrio termodinamico nel passato dell’Universo. Lo studio delle anisotropie permette di selezionare tra diversi modelli per la formazione di strutture quali galassie e ammassi di galassie. Infatti, queste si sono evolute, per instabilità gravitazionale, da piccole perturbazioni di densità all’epoca del disaccoppiamento. Meccanismi quali il redshift gravitazionale e l’effetto Doppler generano delle fluttuazioni della temperatura dei fotoni di fondo cosmico in corrispondenza delle perturbazioni di densità. Quindi la mappa del fondo cosmico permette di ‘fotografare’ la distribuzione iniziale delle fluttuazioni di densità che genereranno successivamente le strutture osservabili nell’Universo.
R. di frenamento
R. elettromagnetica, comunemente detta anche, con termine d’uso internazionale, bremsstrahlung, emessa da un corpuscolo carico il quale subisca, per un motivo qualsiasi, un frenamento. Un caso tipico, e di grande interesse, è quello di un fascio di elettroni interagenti con i campi elettrici dei nuclei degli atomi di un mezzo materiale sul quale il fascio medesimo incida o nel quale si propaghi, utilizzato nei betatroni e negli elettrosincrotroni per la produzione di intensi fasci di raggi γ d’alta energia.
Secondo l’elettromagnetismo classico, un corpuscolo carico, in moto accelerato, irraggia una r. di ampiezza proporzionale all’accelerazione; nel caso di un corpuscolo, di massa m e carica ze (con e valore assoluto della carica dell’elettrone), interagente con il campo di un nucleo atomico, di carica Ze, si trova che, a parità di ogni altra condizione (velocità iniziale del corpuscolo, distanza dal nucleo ecc.), l’accelerazione è proporzionale a Zze2/m e quindi l’intensità della r. è proporzionale a Z2z2e4/m2. L’intensità della r. di frenamento nel caso di un fascio di particelle cariche interagenti con un mezzo materiale viene così a dipendere dal quadrato del numero atomico del materiale attraversato dalle particelle e dall’inverso del quadrato della massa delle particelle medesime: un elettrone irradia circa 106 volte più di un protone e quindi la r. di frenamento degli elettroni è la più rilevante.
R. atmosferiche
Sono il complesso delle r. elettromagnetiche di varia lunghezza d’onda presenti nell’atmosfera terrestre e capaci di influire, soprattutto attraverso i loro effetti termici, sui fattori meteorologici e climatici. Tali r. sono essenzialmente costituite dalle r. inviate dal Sole sulla superficie terrestre (r. solare) e, in misura minore, da quelle che si originano nell’atmosfera (r. del cielo) e al suolo (r. del suolo) in conseguenza dell’assorbimento della r. solare.
R. solare
Le r. elettromagnetiche emesse dal Sole hanno una distribuzione, in funzione della lunghezza d’onda, approssimativamente uguale a quella di un corpo nero alla temperatura di 6000 K e quindi concentrata tra 200 e 1000 nm, nella parte dello spettro per cui l’occhio ha la massima sensibilità. La luminosità del Sole, cioè la potenza totale emessa da tutta la sua superficie, vale 3,85×1026 W. L’intensità della r. alla distanza della Terra e al di fuori della sua atmosfera vale 1367 W/m2, equivalente a circa 8,4 joule (2 calorie) al minuto su ogni cm2 di superficie esposta normalmente ai raggi solari: tale quantità è detta costante solare anche se le misure più recenti, effettuate a bordo di veicoli spaziali, indicano che essa è soggetta a piccole variazioni (dell’ordine di 0,1%) nel tempo.
Le r. che giungono al suolo sono attenuate e in alcune lunghezze d’onda anche estinte da parte dell’atmosfera terrestre, in seguito a fenomeni di riflessione, diffusione e assorbimento. Infatti, una parte della r. solare viene riflessa verso gli spazi cosmici da molecole, polveri e nubi di vapori contenuti nell’atmosfera; in particolare le r. radio di grande lunghezza d’onda (all’incirca maggiore di 20 m), avendo una frequenza minore della frequenza critica della ionosfera, sono completamente riflesse da questa. Una certa parte della r. non riflessa è poi diffusa in tutte le direzioni dalle molecole dei gas atmosferici, costituendo la r. diffusa. Poiché la diffusione avviene con un’intensità che è in ragione inversa della quarta potenza della lunghezza d’onda (➔ diffusione), la r. diffusa è costituita dalle r. solari di breve lunghezza d’onda, con un massimo d’intensità all’incirca per l’azzurro dello spettro visibile (è appunto la r. diffusa che determina il colore azzurro del cielo); conseguentemente, lo spettro della r. non diffusa (r. diretta o trasmessa) risulta attenuato nel campo delle r. di più breve lunghezza d’onda. Ma il processo di gran lunga più importante è l’assorbimento che la r. solare, sia quella diffusa sia quella diretta, subisce nell’atmosfera.
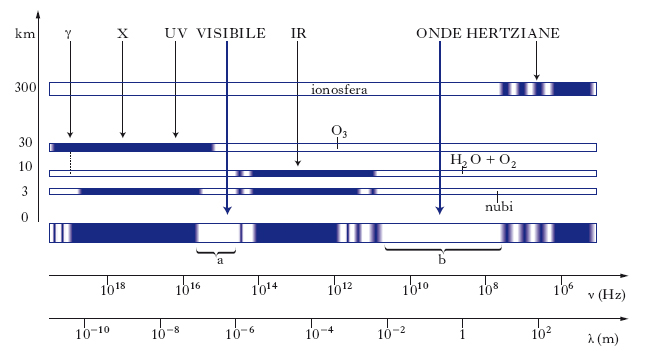
I risultati di numerose misurazioni hanno mostrato che l’assorbimento atmosferico è fortemente selettivo: l’atmosfera è in pratica trasparente solo in due regioni dello spettro che, con locuzione molto espressiva, sono dette ‘finestre dell’atmosfera’ (fig. 4). La prima (finestra ottica), estendentesi da circa 0,1 a circa 10 μm di lunghezza d’onda, comprende una piccola parte delle r. ultraviolette, l’intero spettro visibile e una piccola parte delle r. infrarosse; la seconda (finestra radio) comprende le onde hertziane fra circa 1 cm e circa 30 m di lunghezza d’onda. In particolare sono quasi completamente estinte le r. a brevissima lunghezza d’onda e alta energia (raggi γ, raggi X e, in parte, i raggi ultravioletti): tale fatto, fondamentale per il mantenimento sulla Terra della vita nelle sue attuali forme, è dovuto alla presenza nell’atmosfera dell’ozono e, in minor misura, dell’ossigeno, che assorbono tali radiazioni. L’assorbimento delle r. appartenenti alla zona dello spettro compreso tra le due finestre e quindi, in particolare, della maggior parte delle r. infrarosse, è dovuto essenzialmente al vapore acqueo e all’ossigeno.
Anche il suolo terrestre contribuisce alla riflessione e all’assorbimento delle r. solari. La Terra, nel suo complesso, cioè superficie terrestre e atmosfera, riflette in media il 45% delle r. solari incidenti. La superficie terrestre assorbe il 75% di tutta la r. solare assorbita, mentre l’atmosfera ne assorbe solo il 25%: il suolo, reirradiando energia, riscalda a sua volta sensibilmente l’atmosfera. Per il valore dell’intensità della r. solare globale (diretta e diffusa) al suolo, si hanno ampie variazioni nel corso della giornata e dell’anno, in dipendenza del variare dell’altezza del Sole sull’orizzonte (e quindi dello spessore dello strato d’aria attraversato) e in dipendenza delle condizioni atmosferiche (nuvolosità, stato igrometrico, presenza di pulviscolo ecc., fattori tutti che alterano la trasparenza dell’atmosfera). Tale intensità è in media dell’ordine di 700 W/m2 nelle ore meridiane di una giornata estiva con cielo sereno e atmosfera limpida: l’atmosfera, cioè, nelle migliori condizioni di trasparenza, assorbe circa la metà della r. solare.
La r. corpuscolare solare è costituita da una componente stabile (un plasma di bassa energia detto vento solare) e da una componente impulsiva, legata ai brillamenti, formata da protoni e altri nuclei atomici con energie da alcuni MeV a oltre 103 MeV. La prima componente è bloccata dal campo geomagnetico a grandi distanze dalla Terra; la seconda raggiunge il suolo, ma non contribuisce in modo apprezzabile al riscaldamento atmosferico.
R. termica del cielo e del suolo
L’assorbimento della r. solare determina un certo riscaldamento dell’atmosfera e del suolo; si originano così due r. secondarie, entrambe di natura termica, cioè a relativamente grande lunghezza d’onda (con massimo d’intensità circa sui 10 μm): una diretta dal suolo all’atmosfera (r. del suolo), l’altra, originantesi nei bassi strati atmosferici, diretta quasi esclusivamente verso il suolo (r. del cielo o controradiazione atmosferica). L’esistenza di queste due r. provoca un certo flusso di calore (r. termica suolo-atmosfera), diretto, a seconda del prevalere dell’una o dell’altra, dal suolo all’atmosfera o viceversa. Poiché l’intensità della r. suolo-atmosfera è calcolabile a mezzo della legge di Stefan (➔ corpo) nota che sia la temperatura termodinamica del suolo, le misurazioni con il pirgeometro permettono di risalire all’intensità della r. del cielo. Dal valore di questa si risale poi, sempre a mezzo della legge di Stefan, alla temperatura virtuale del cielo, cioè alla temperatura alla quale un corpo nero emetterebbe le r. emesse dall’atmosfera. L’intensità della r. suolo-atmosfera è, naturalmente, molto minore di quella della r. solare, raggiungendo, al massimo, circa 150 W/m2; di giorno è diretta verso il suolo, di notte verso l’atmosfera, raggiungendo il massimo valore poco prima dell’alba. La r. suolo-atmosfera notturna, detta brevemente r. notturna, ha una notevole importanza meteorologica: è infatti la r. notturna che, provocando un più o meno sensibile raffreddamento del suolo, determina la formazione della rugiada e della brina.
Biologia
La disciplina che studia gli effetti delle r. sui materiali biologici è la radiobiologia (➔) e radiobiologici si denominano i fenomeni da essa contemplati che riguardano gli esseri viventi. Tutti questi fenomeni sono conseguenza dell’assorbimento di energia ceduta dal fascio di r. nel materiale irradiato. La quantità di energia dissipata per unità di massa del materiale rappresenta la dose fisica di r. assorbita dal materiale stesso e viene misurata in gray (simbolo Gy; pari a 1 J/kg), sostituito al rad (pari a 10–2 Gy).
Effetti sugli acidi nucleici
La struttura primaria del DNA è costituita da uno ‘scheletro’ di fosfati alternati con gruppi di pentosi, a cui è legata una base azotata purinica (G, C) o pirimidinica (A, T; ➔ nucleici, acidi). Il danno indotto dalle r. coinvolge principalmente le basi pirimidiniche. Bisogna distinguere due effetti, a seconda che le r. inducano eccitazione elettronica o ionizzazione. Alla prima categoria appartengono le r. ultraviolette che, a una lunghezza d’onda di 260 nm, determinano dimerizzazione di due timine adiacenti sullo stesso filamento di DNA e provocano la formazione di legami covalenti o anelli di ciclobutano. Le r. ionizzanti, invece, attraverso processi legati alla presenza di radicali liberi OH•, determinano il distacco e l’asportazione delle basi con la formazione di siti privi di purine (apurinici) o privi di pirimidine (apirimidinici).
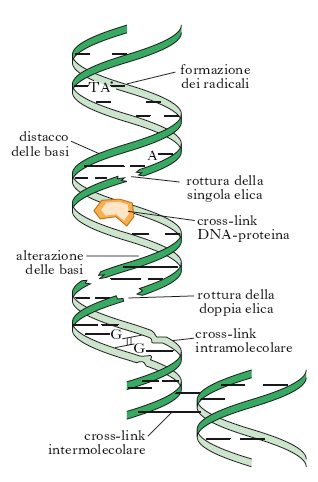
Altri tipi di lesioni indotte dalle r. ionizzanti nella struttura primaria del DNA sono: rotture della singola elica (single strand break, SSB), rotture della doppia elica (double strand break, DSB) e legami crociati fra le due eliche del DNA o fra DNA e proteine (fig. 5). Le rotture a singolo filamento avvengono in seguito all’idrolisi del legame fosfodiesterico e all’eliminazione di un gruppo fosfato, con un meccanismo che coinvolge i radicali liberi dell’acqua. È stato dimostrato che il numero di SSB è correlato con la dose di r. somministrate. L’energia necessaria per la formazione di rotture a doppio filamento è di 1000-1800 eV. L’interazione tra radicali radioindotti può dare luogo, anche se con bassa frequenza, a legami tra eliche di due molecole distinte, o tra una molecola di DNA e una molecola proteica (cross-link).
Effetti sulle proteine
L’azione indiretta delle r. coinvolge proteine con funzioni sia strutturali sia enzimatiche, alterando la sequenza amminoacidica e la configurazione spaziale. Per produrre una inattivazione apprezzabile di un enzima in vivo sono necessarie decine di Gy, mentre solo 2 Gy sono sufficienti per indurre la morte di una popolazione cellulare.
Effetti sulle membrane
I radicali liberi prodotti dalle r. possono anche interagire con le membrane plasmatiche attraverso processi di perossidazione dei lipidi (➔ perossidi) e di ossidazione dei gruppi sulfidrilici delle proteine. In generale si può dire che le r. inducono sulle membrane cellulari una riduzione della loro fluidità: si verifica una modificazione della permeabilità agli ioni a basse dosi (1-50 Gy) o un aumento della permeabilità passiva ad alte dosi (100-200 Gy). Ad alte dosi di r. è stata inoltre osservata un’alterata permeabilità cellulare nei lieviti, negli eritrociti e in cellule muscolari, mentre una modificazione dell’ampiezza degli impulsi nervosi è stata osservata nei nervi periferici isolati.
Effetti sulle cellule
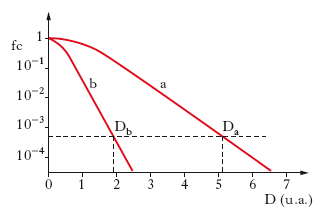
La capacità riproduttiva delle cellule sia in vivo sia in vitro costituisce un buon parametro per misurare l’effetto delle r. sul materiale vivente. Si possono distinguere due fenomeni: la morte cellulare e quella riproduttiva. Si parla di morte cellulare quando cellule non proliferanti, quali nervi e muscoli, perdono la loro funzione biologica; si parla invece di morte riproduttiva quando cellule attivamente proliferanti, quali cellule staminali, ematopoietiche o cellule coltivate in vitro, perdono la capacità di riprodursi. La capacità di una singola cellula di riprodursi e formare una colonia è una prova della sua integrità. La perdita di tale funzione in relazione alla dose di r. è quantitativamente descritta dalle cosiddette curve di sopravvivenza delle popolazioni irradiate, che esprimono la percentuale di cellule sopravviventi (in ordinata logaritmica) in funzione della dose somministrata (in ascissa lineare): un esempio schematico per irradiazione acuta con raggi X o γ è dato nella curva a della fig. 6. La curva presenta, alle basse dosi, una ‘spalla’ iniziale seguita, per dosi progressivamente crescenti, da un tratto finale con andamento approssimativamente esponenziale. La spalla iniziale è in rapporto con la necessità che la cellula accumuli, nei cromosomi, una certa quantità di danno, prima che la sopravvivenza si riduca di un valore costante a unità di dose. La dose necessaria per ridurre del 37% le cellule sopravviventi nel tratto esponenziale della curva è denominata Da e caratterizza, per convenzione, la radiosensibilità intrinseca della popolazione cellulare. Poiché cellule in fasi diverse del ciclo di divisione hanno sensibilità diversa, la curva di sopravvivenza di una popolazione in accrescimento esponenziale è la media risultante dalla risposta delle varie sottopopolazioni. Altri numerosi fattori di natura fisica, fisico-chimica o chimica possono variamente modificare la forma delle curve di sopravvivenza. Cellule irradiate con particelle ad alta densità lineare di ionizzazione (neutroni, r. α) mostrano curve di sopravvivenza con una parte iniziale più ridotta e un tratto esponenziale più ripido, come nella curva b della fig. 6; esse risultano cioè più efficaci nell’indurre l’effetto. Il rapporto Da/Db tra la dose della r. standard e quella di confronto per produrre un certo livello di effetto rappresenta l’efficacia biologica relativa (EBR) di una r. rispetto all’altra. Quest’effetto non è in relazione con la quantità di energia depositata per unità di massa del tessuto irradiato, ma con il modo in cui l’energia viene depositata dai vari tipi di r., cioè con la densità lineare di ionizzazione.

Lo studio degli effetti delle r. in cellule di mammifero coltivate in vitro ha permesso di conoscere la relazione fra r., ciclo cellulare e riparazione. Le varie fasi del ciclo cellulare sono strettamente condizionate dal completamento della fase precedente. La successione delle varie fasi del ciclo è regolata da specifici punti di controllo (checkpoints). Dopo esposizione a r. le cellule sono in grado di rallentare drasticamente la loro progressione, accumulandosi in punti di controllo tra la fase G1 e S (G1/S) e la fase G2 e M (G2/M) del ciclo cellulare. Si determina in tal modo rispettivamente un blocco in G1 e/o in G2. Questi rallentamenti transitori concedono un tempo maggiore alle attività riparative, limitando il rischio di avere, nella fase di sintesi del DNA (fase S) e quindi nelle generazioni successive, cellule fortemente danneggiate. La radiosensibilità nelle diverse fasi del ciclo cellulare è stata osservata utilizzando cellule sincronizzate (tutte nella stessa fase del ciclo cellulare). Le fasi M e G2 sono le più sensibili, mentre più radioresistenti sono le cellule in fase S (fig. 7). Secondo alcuni autori queste variazioni possono essere messe in relazione con la conformazione e la compattezza del DNA nelle diverse fasi del ciclo oppure con la presenza quantitativamente maggiore di gruppi sulfidrilici in fase S rispetto alla mitosi e alla G1.
Effetti sulla struttura del cromosoma
L’effetto delle r. è stato largamente studiato osservando la struttura dei cromosomi nella prima metafase dopo il trattamento in cellule somatiche attivamente proliferanti (meristemi di radici di Vicia faba, cellule in coltura, linfociti di sangue periferico). Dato che tutti i tipi di r. sono in grado di indurre lesioni al DNA, lesioni mal riparate o non riparate sono la causa di alterazioni strutturali dei cromosomi chiamate aberrazioni cromosomiche (➔ mutazione). Successivi studi hanno messo in evidenza che le lesioni responsabili della formazione delle aberrazioni cromosomiche sono le rotture della doppia elica del DNA.
I diversi tipi di aberrazioni possono essere classificati come aberrazioni di tipo cromatidico o cromosomico, a seconda se sono coinvolti nel danno uno o ambedue i cromatidi di uno stesso cromosoma. Un altro tipo di classificazione prende in considerazione la possibilità che l’alterazione della struttura dei cromosomi possa essere trasmessa nelle cellule figlie, e in questo caso si parlerà di aberrazioni stabili, quali traslocazioni, o aberrazioni instabili, quali dicentrici (scambi fra cromosomi con perdita di frammenti acentrici). Nelle traslocazioni si ha la conservazione di tutto il materiale genetico, mentre i dicentrici costituiscono degli impedimenti alla regolare divisione mitotica e sono una delle cause della morte cellulare.
Riparazione del danno da radiazioni
Ogni effetto radiobiologico può essere considerato come la risultante di due forze che operano in senso opposto: da una parte, il danno indotto dalla r. sulle strutture sensibili della cellula; dall’altra, i meccanismi che ogni sistema biologico può spontaneamente mettere in atto per riparare tale danno. Gli effetti della riparazione del danno vengono studiati sia a livello molecolare (➔ riparazione) sia a livello cellulare. Diversi tipi di riparazione sono stati descritti in cellule di mammifero in coltura e verificati poi in modelli sperimentali in vivo. In assenza di divisione cellulare e in condizioni sperimentali adatte, si verifica una riparazione del cosiddetto danno subletale. In questo caso le cellule, durante l’intervallo tra le due dosi somministrate nell’arco di alcune ore, riparano il danno inflitto dalla prima. Il danno, dopo due dosi intervallate, risulterà quindi inferiore a quello atteso dopo la somministrazione di una singola irradiazione di intensità uguale alla somma delle due. Un altro tipo di riparazione riguarda il danno potenzialmente letale: quando si ritarda sperimentalmente l’inizio della divisione cellulare successiva all’irradiazione, il risultato è che il numero di cellule inattivate risulta inferiore a quello che ci si sarebbe atteso se le cellule fossero state libere di dividersi normalmente.

La riparazione del danno subletale e di quello potenzialmente letale sono particolarmente evidenti dopo irradiazione X o γ. Esiste infine un tipo diverso di riparazione, che riguarda l’intero tessuto: si realizza quando la frazione di cellule che sopravvive a una certa dose, dopo un iniziale blocco della divisione ricomincia a proliferare, ripopola il tessuto irradiato e ne ricostituisce così l’integrità. I fenomeni di riparazione descritti spiegano un’osservazione del tutto generale, cioè che riducendo l’intensità della dose si ottiene di solito una riduzione dell’effetto. Una bassa dose somministrata cronicamente si può infatti considerare come una successione di piccole frazioni, e si può pertanto comprendere come la curva di sopravvivenza diventi progressivamente meno ripida, come in fig. 8. Ciò avviene, in un primo tempo, in conseguenza della riparazione del danno e poi a seguito della proliferazione delle cellule sopravviventi.
Gli effetti delle r. sull’uomo
Gli effetti delle r. nei tessuti e nell’organismo umano possono essere la conseguenza o della morte di un gran numero di cellule nel tessuto (effetto somatico) o di danni nell’embrione (effetto teratogeno). Manifestazioni del danno in una singola cellula possono essere la causa del cancro (effetti stocastici) o avere conseguenze nelle generazioni successive (effetti genetici).
Effetti somatici
La sensibilità di un tessuto dipende dal grado di differenziazione e dalla capacità proliferativa delle cellule che lo costituiscono. La dose di r. necessaria per distruggere una funzione cellulare in cellule differenziate è molto più alta di quella necessaria per bloccare l’attiva divisione cellulare. I danni si riscontrano a livello del midollo osseo (leucopenia, linfopenia, piastrinopenia, anemia aplastica), dell’intestino tenue (perdita dell’epitelio, ulcerazione, alterazione dell’equilibrio idrico-salino), della cute e dei suoi annessi (eritema, desquamazione secca e umida, ulcerazione, necrosi, depilazione), degli organi maschili della riproduzione (sterilità transitoria o permanente). Tutti questi tessuti sono strutturati per compartimenti gerarchici, a rapido rinnovamento, e hanno quindi una patogenesi analoga, anche se i sintomi clinici e i tempi di insorgenza sono diversi. I tessuti a rinnovamento più lento, quali il fegato, il polmone, il rene, che hanno una struttura funzionale flessibile, hanno anche risposte più ritardate e sono in genere sensibili soltanto a dosi relativamente elevate.
L’intervallo di tempo fra l’irradiazione e l’espressione del danno nel tessuto irradiato dipende dall’intervallo di tempo necessario perché una cellula da una fase indifferenziata (cellula staminale) si differenzi, acquistando tutte le funzioni fisiologiche tipiche del tessuto. Cellule staminali che proliferano attivamente, quali le cellule delle cripte intestinali, esprimeranno il danno dopo un breve intervallo di tempo. L’esposizione a r. di cellule staminali del sangue circolante avrà, invece, quale prima conseguenza, la riduzione delle cellule in questo tessuto, e solo successivamente la manifestazione del danno.
In un organismo esposto a r. è necessario distinguere se l’esposizione è stata limitata soltanto a una parte del corpo o è stata totale: nel primo caso le conseguenze riguardano principalmente il tessuto o l’organo irradiato; nel secondo caso si parlerà di ‘panirradiazione’ e le osservazioni verranno effettuate su tutti i tessuti separatamente e contemporaneamente. Individui esposti a irradiazione mostrano: a) una sindrome neurologica, caratterizzata da gravi lesioni a carico del sistema nervoso centrale e da profondo stato di shock, che si osserva entro poche ore o pochissimi giorni da un trattamento acuto su tutto il corpo con raggi X o γ (dosi dell’ordine di alcune decine di Gy); b) una sindrome intestinale, che si manifesta entro alcuni giorni o un paio di settimane con vomito, diarrea, emorragie, febbre (10 Gy o più); c) una sindrome ematologica, che può comparire, dopo l’irradiazione, in un periodo che va da pochi giorni fino a due mesi, con infezioni, febbre, emorragie, perdita di peli e capelli (2-10 Gy).
Si ritiene che la dose letale per il 50% degli individui irradiati, nel giro di circa due mesi e in assenza di qualsiasi terapia, sia tra 3 e 6 Gy. La cura della sindrome neurologica e intestinale può essere solo sintomatica e palliativa, essendo esse invariabilmente letali; quella della sindrome ematologica consiste unicamente nel trapianto del midollo osseo.
Effetti teratogeni
Gli effetti teratogeni delle r. nei vari stadi embrionali e fetali dello sviluppo sono stati osservati in animali da laboratorio, ma le conclusioni raggiunte sono applicabili anche all’uomo. La fase che precede l’impianto dell’ovulo fecondato è quella più sensibile; in questa fase le r. causano la morte prenatale dell’embrione; nella fase successiva all’impianto dell’embrione nell’utero e nella organogenesi le conseguenze delle r. sono, invece, ritardo nella crescita e malformazioni dell’embrione. È stato valutato che una dose di 0,05 Gy di r. induce una letalità prenatale compresa fra 0 e 1 ogni 1000 nati; un analogo rischio si può osservare anche nell’insorgenza di gravi alterazioni del sistema nervoso centrale con conseguente ritardo mentale.
Effetti stocastici
Riguardano soprattutto la relazione fra r. e cancro. Per questo tipo di fenomeno non si può parlare di dose soglia, dal momento che anche il danno di una sola cellula può determinare gravi conseguenze nell’individuo; alla base di questa considerazione vi è l’origine clonale del cancro. Lo sviluppo di neoplasie come effetto tardivo delle r. e le cause che le determinano sono stati oggetto di numerosi studi sin dall’inizio del 20° sec. e la valutazione del rischio di sviluppo di leucemie o di altri tipi di tumori è stata effettuata mediante studi epidemiologici in popolazioni umane. Anche se il meccanismo non è completamente chiarito, la teoria più accreditata sull’origine dei tumori è attualmente quella della mutazione somatica e dell’attivazione e dell’espressione di oncogeni (➔ oncogene; protooncogene). L’intervallo fra l’irradiazione e la comparsa di cellule tumorali può durare anni e decenni. La cancerogenesi è un processo complesso a più stadi: una fase di inizio in cui la cellula viene danneggiata, una fase di promozione quando la cellula inizierà a dividersi, e una fase di progressione legata alla capacità delle cellule stesse di crescere in maniera incontrollata.
Il periodo che intercorre fra la r. e lo sviluppo del cancro viene definito ‘tempo di latenza’ e può variare in funzione dell’età dell’individuo irradiato e del tipo di cellule coinvolte, ma non in funzione della dose. Dati ottenuti su abitanti di Hiroshima e Nagasaki, sottoposti alle radiazioni della bomba atomica nel 1945, hanno messo in evidenza che la leucemia è il tipo di tumore con minore periodo di latenza: infatti si manifesta già dopo 7-12 anni dall’esposizione e la sua frequenza decresce fino a raggiungere i valori normali caratteristici della popolazione dopo 20 anni: al contrario, i tumori solidi si manifestano dopo 10-20 anni dall’irradiazione dell’individuo. Il tempo di latenza varia a seconda dei tipi di tessuto considerati, quali tiroide, seno, polmone, stomaco ecc. Inoltre, in una popolazione non è facile distinguere la percentuale di tumori attribuibile alle r. da quella attribuibile a virus o a fattori inquinanti ambientali. I dati che mettono in relazione le r. e il cancro derivano quasi tutti da esposizioni ad alte dosi dovute a incidenti, mentre pochi sono i dati a disposizione sulle basse dosi, che sono quelle di maggiore interesse per la popolazione.
Effetti genetici
Si definiscono effetti genetici i cambiamenti indotti dalle r. nel materiale ereditario dei gameti di un individuo e che possono venire trasmessi alle generazioni successive. Le mutazioni possono essere dominanti se compaiono già nella prima generazione dopo l’irradiazione, recessive se si manifestano solo quando due alleli portatori della stessa mutazione, provenienti dai due genitori, si trovano ambedue presenti nel genotipo dello zigote. Le mutazioni possono interessare sia caratteristiche morfologiche sia processi essenziali alla vita, il cui cambiamento determina la morte dell’individuo (mutazioni letali). Un aumento della frequenza di mutazioni causato da r. fu dimostrato per la prima volta nel 1927 da H.J. Miller in cellule germinali di Drosophila melanogaster. Avvalendosi di una complessa serie di incroci, egli studiò la frequenza di mutazioni letali recessive indotte dalle r. a basso LET (linear energy transfer) sul cromosoma X. I risultati evidenziarono che la frequenza di mutazione aumentava linearmente con la dose della radiazione. Un parametro da tenere in considerazione è la dose di raddoppio, che corrisponde alla dose di r. necessaria per raddoppiare la frequenza di mutazioni spontanee di un determinato gene. In tutte le specie studiate è stato sempre osservato, dopo irradiazione, un aumento delle mutazioni. Questo si riscontrò anche nel topo, il primo mammifero studiato, le cui cellule germinali possiedono una radiosensibilità otto volte superiore a quella di Drosophila. Oltre alle mutazioni geniche le r. possono indurre nei gameti alterazioni strutturali dei cromosomi; in tal caso l’individuo portatore di traslocazione sarà sterile o semisterile oppure potrà dare origine a individui malformati. Non solo le r. ionizzanti ma anche le r. ultraviolette a una lunghezza d’onda di 260 nm sono in grado di indurre mutazioni che si esprimono nella progenie. Molti studi sono stati condotti su virus, Batteri, polline di piante superiori, ma non vi sono dati sui Mammiferi a causa del basso potere di penetrazione delle r. ultraviolette nei tessuti profondi.
Pochi sono i dati sugli effetti genetici delle r. nell’uomo. Gli unici gruppi studiati a questo riguardo appartengono alle popolazioni sopravvissute all’esplosione della bomba atomica. In queste popolazioni sono stati considerati quattro parametri: a) gli aborti e le malformazioni congenite; b) la morte nei primi anni di vita; c) le anomalie dei cromosomi sessuali; d) le variazioni elettroforetiche di proteine. I risultati sono comunque di difficile interpretazione per vari ordini di motivi, quali la mancanza di controlli adeguati e il fatto che le mutazioni recessive si possono manifestare anche dopo molte generazioni. Solo ora, infatti, cominciano a essere pubblicati i dati riguardanti la mobilità elettroforetica delle proteine e le alterazioni strutturali dei cromosomi, nella prima generazione nata dopo l’esplosione delle bombe atomiche nel 1945. Comunque non vi sono finora motivi per far credere che i risultati ottenuti su altri Mammiferi, quali il topo, non siano validi anche per l’uomo. A tale proposito è consigliato a individui esposti ad alte dosi di r. un periodo di 6 mesi prima di programmare un concepimento, per ridurre il rischio di anomalie genetiche.
Tutti gli effetti biologici delle r. descritti possono essere amplificati dalla presenza di ossigeno nelle cellule o nei tessuti irradiati; al contrario, agenti chimici con gruppi sulfidrilici SH vengono considerati dei radioprotettori.
Effetti biologici delle r. non ionizzanti
Gli effetti prodotti dalle r. elettromagnetiche di lunghezza d’onda maggiore di circa 5∙10–7 m, cioè comprese tra la luce visibile e i campi statici, se si prescinde da quelli puramente termici, non sono riconducibili a un unico meccanismo d’azione e il loro studio è in rapida evoluzione. La configurazione elettronica di numerose biomolecole è tale che fotoni nel visibile possono indurre transizioni tra due diversi livelli elettronici: lo stato eccitato, se ha una vita media sufficientemente lunga ed è chimicamente attivo, può innescare processi di profonda rilevanza per il metabolismo cellulare (si può, per es., avere la formazione di composti letali per la cellula). L’esposizione di sistemi biologici a onde millimetriche sembra mostrare l’esistenza di fenomeni di risonanza tra le onde stesse e domini di dipoli elettrici in membrane biologiche. Si hanno infine molte evidenze sperimentali di modificazioni in vitro delle funzioni cellulari, in particolare di cellule di origine nervosa, immunologica e osteoblastica, ottenute con r. elettromagnetiche di lunghezza d’onda maggiore di circa 10 m (cioè di frequenza inferiore a circa 30 MHz) che produrrebbero la variazione della concentrazione dello ione Ca++ intracellulare; i dati in vitro sono affiancati da evidenze cliniche come la terapia elettromagnetica di fratture ossee che non consolidano spontaneamente.
© Istituto della Enciclopedia Italiana - Riproduzione riservata


