Recettori
Enciclopedia del Novecento (1982)
Recettori
di Ragnar Granit
SOMMARIO: 1. Concetti generali: a) cenni storici; b) tipi di recettori; c) definizioni elementari; d) potenziale generatore; e) adattamento; f) altri modi di trasmettere informazioni sensoriali; g) campi recettivi; h) importanza della dimensione della fibra; i) soglie. □ 2. Recettori specifici: a) il corpuscolo di Pacini; b) il fuso neuromuscolare dei Mammiferi; c) recettori cutanei; d) recettori olfattivi; e) controllo inibitorio centrifugo; f) linea laterale, apparato vestibolare. □ Bibliografia.
1. Concetti generali.
a) Cenni storici.
Lo studio dei recettori nel senso moderno cominciò nella prima metà del secolo scorso come una scienza delle correlazioni fra stimolo e risposta, chiamata poi psicofisica (v. Fechner, 1862). La percezione sensoriale conscia era definita dalle proprietà fisiche dello stimolo. Alcuni tipi di stimoli non si potevano definire molto accuratamente; per esempio, le deformazioni della pelle che producono forze agenti sui recettori cutanei si potevano approssimare solo in grammi di peso: si doveva supporre che le tensioni prodotte dalla pressione fossero proporzionali al peso applicato. La psicofisica ottenne i suoi risultati migliori con i telecettori come l'occhio e l'orecchio per i quali era possibile graduare accuratamente gli stimoli. La lunghezza d'onda in uno spettro poteva definire i colori; in acustica il suo valore inverso, la frequenza, definiva i toni ecc. La ‛soglia' ovvero il ‛minimo percettibile' e il periodo latente della risposta erano altre quantità accessibili allo studio con metodi psicofisici. L'intensità dello stimolo era messa in relazione all'‛intensità' percepita, misurabile come luminosità nella visione e come intensità del suono in acustica, e questa relazione venne definita dalla legge di WeberFechner che stabilisce che quando l'intensità dello stimolo aumenta in progressione geometrica, qualcosa nella sensazione, che possiamo chiamare la sua quantità o intensità, aumenta in progressione aritmetica.
Tuttavia, in psicofisica il rivelatore era necessariamente psicologico e cosi il contributo dell'organo terminale non si poteva mai valutare inequivocabilmente. Lo studio oggettivo delle proprietà dei recettori dovette attendere l'introduzione in fisiologia dell'elettronica, all'inizio degli anni venti. Amplificatori e strumenti di registrazione diretta, quali l'oscillografo a raggi catodici, resero possibile registrare gli impulsi in singole fibre di nervi afferenti (v. Adrian, 1928 e 1932) e infine l'attuale tecnica dei microelettrodi extrae intracellulari ha sottoposto all'osservazione diretta gli stessi organi terminali. Le conquiste dell'ultimo mezzo secolo di ricerca sui recettori saranno l'argomento di questa rassegna, ma è giusto riconoscere che la scienza della psicofisica ha creato un solido fondamento a tutto il campo, in particolare per i cosiddetti ‛sensi superiori' (occhio, orecchio) che dominano la nostra esperienza cosciente. Riguardo agli organi vestibolari dell'orecchio interno, che nella posizione del corpo e nel movimento consentono l'orientamento secondo la gravità, studi comportamentali (v. Flourens, 1830), associati a distruzioni o stimolazioni localizzate di tali organi, dettero buoni indirizzi per ulteriori sviluppi (v. Granit, 1955; v. von Kries, 1923).
b) Tipi di recettori.
Gli organi di senso si possono definire gli strumenti di misura dell'organismo vivente e come tali essi hanno delle proprietà come sensibilità, ambito di lavoro, velocità e potere di risoluzione (v. Granit, 1955). Negli animali in movimento essi raggiungono il loro più alto grado di perfezione; questo appare facilmente comprensibile, perché movimenti dettati, per esempio dalla paura, dalla fame o dal desiderio sessuale sono atti finalistici richiedenti informazioni che ovviamente sono basate sulla lettura di questi strumenti, specialmente dei telecettori della vista, dell'udito e dell'olfatto. Gli stessi movimenti per essere eseguiti armoniosamente devono essere controllati dalle informazioni sensitive provenienti dagli organi terminali dei muscoli, dei legamenti e delle articolazioni. Questo è noto da molto tempo e uno dei più vecchi esperimenti di neurologia dimostra che nei Mammiferi l'interruzione delle informazioni in arrivo, realizzata mediante sezione delle radici dorsali afferenti del midollo spinale, provoca gravi deficienze nell'esecuzione dei movimenti.
Mentre noi siamo consci dei messaggi provenienti dai sensi che interpretano il nostro ambiente, v'è un gran numero di organi terminali che agiscono, per così dire, in segreto, ed è stato necessario molto lavoro sperimentale per chiarire che cosa essi misurano e in che modo. A questa categoria appartengono gli strumenti che guidano gli atti motori misurando automaticamente la lunghezza e la tensione dei muscoli che si contraggono, e le terminazioni delle articolazioni sensibili allo spostamento: questi organi sono chiamati propriocettori. Analogamente, non siamo consci di ciò che fanno molti enterocettori, quali, per esempio, quelli stimolati dalle variazioni della tensione polmonare, della pressione o della composizione chimica del sangue. Ciò nonostante questi sensi sono organi di controllo importantissimi, i cui effetti fisiologici possono essere studiati in modi molto diversi. La nostra esperienza cosciente è dominata dagli esterocettori, le cui proprietà di misura sono adatte agli stimoli provenienti dal mondo esterno. Fra questi predominano i telecettori, che sono generalmente situati nella testa dell'animale.
La necessità di informazioni di un organismo varia a seconda del suo tipo di vita, e animali che volano o nuotano hanno organi terminali adattati alle condizioni prevalenti rispettivamente nell'aria o nell'acqua. I pipistrelli, per es., hanno orecchi sensibili agli ultrasuoni (suoni ad alta frequenza) e hanno organi vocali capaci di emettere suoni alle corrispondenti frequenze, che, riflessi dagli oggetti circostanti, servono a guidarli nel loro volo a mo' di radar. I Pesci hanno una linea laterale di organi di senso che controllano il loro movimento nell'acqua (v. cap. 2, È f). Alcune specie di Pesci, capaci di generare nell'acqua circostante un campo elettrico di corrente oscillante, hanno speciali elettrocettori che misurano la resistenza che questa corrente incontra a causa degli oggetti presenti nell'interno del campo; alcuni di questi pesci vivono in ambienti in cui l'acqua tende a essere fangosa, rendendo così la vista meno utile che in condizioni normali. Negli Invertebrati si sta scoprendo un numero sempre crescente di nuovi e spesso sorprendenti organi di senso, strumenti capaci di rivelare il piano di polarizzazione della luce (studiati esaurientemente nelle api: v. von Frisch, 1950), recettori igroscopici, rivelatori di raggi infrarossi e ultravioletti, per citare alcuni esempi.
c) Definizioni elementari.
Lo stimolo ‛adeguato' o specifico di un qualsiasi organo terminale dei Mammiferi tende a essere unico: luce, suono, freddo, caldo, tatto, ecc., e dal punto di vista di una buona discriminazione questo sembrerebbe un disegno ideale. Si definisce stimolo adeguato quello al quale la terminazione presenta la massima sensibilità o - più tecnicamente - per il quale ha la soglia più bassa. La vecchia dottrina delle ‛energie nervose specifiche' di Müller (v., 1838) asserisce che la specificità dell'organo terminale è determinata dalla corrispondente specificità in una struttura interpretativa centrale: il che significa, per es., che con qualsiasi stimolo inadeguato si stimoli il nervo ottico o acustico (per es., elettrico o meccanico) la sensazione sarà sempre rispettivamente luminosa o sonora. Doppie specificità, però, sia pure limitatamente al freddo e alla pressione, non sono del tutto sconosciute nei Mammiferi, come diremo in seguito (v. cap. 2, È c): la lingua del gatto possiede alcune terminazioni sensibili sia alla pressione sia al freddo (v. Hensel e Zotterman, The response of..., 1951). A un livello inferiore nella scala zoologica, la cavità facciale del serpente a sonagli è sensibile alla radiazione infrarossa e al tatto (v. Bullock e Horridge, 1965) e probabilmente saranno scoperti nuovi esempi negli Invertebrati. Le mosche, per es., hanno chemocettori sensibili anche alla temperatura e agli stimoli meccanici. A che servirebbe l'informazione mista come quella provvista dalle triplici specificità di questo genere? Abituati come siamo all'idea che un organo di senso rappresenti solo una ‛modalità', ci è difficile immaginare che l'emissione delle informazioni sia realmente l'unica chiave per la comprensione delle specificità miste. Gli organi di senso hanno un'altra funzione, quella di servire da attivatori generici o aspecifici dei neuroni ai quali inviano messaggi: questo compito differisce da quello di fornire informazioni sensoriali specifiche e chiaramente è un incentivo al mantenimento come tale dell'attività afferente. Sembrerebbe che la sensibilità a molti tipi di stimoli possa servire a facilitare delle prolungate scariche di impulsi verso quelle stazioni centrali che ne hanno bisogno per mantenersi reattive.
Oltre che per la modalità, la sensazione si distingue anche per la ‛qualità': prendendo come esempio di una modalità la vista, i differenti colori si chiamerebbero qualità visive.
La sensibilità specifica a uno stimolo adeguato implica che la terminazione ricevente di un recettore è capace di reagire selettivamente a una particolare forma di energia e di trasformarla in uno stimolo interno per il sistema vivente. Per questa capacità i recettori sono definiti i ‛trasduttori' biologici di una forma di energia in un'altra, nello stesso modo in cui una membrana telefonica serve da trasduttore dell'energia meccanica in energia elettrica.
In un solo esempio, quello dei bastoncelli della retina, si conosce a grandi linee il processo iniziale specifico a livello molecolare. In questi recettori i quanti di luce sono assorbiti dalla sostanza fotosensibile, la rodopsina (v. visione), la cui scissione e trasformazione in risposta alla stimolazione luminosa sono state studiate dettagliatamente. Poiché la reazione iniziale è di natura chimica, i bastoncelli si possono classificare come chemocettori insieme con gli organi terminali dell'olfatto e del gusto. Un grandissimo gruppo di recettori appartiene alla categoria generica dei meccanocettori, come le terminazioni sensoriali dell'orecchio, quelle sensitive dei fusi neuromuscolari, dei tendini, dei legamenti, delle vibrisse e di molti tipi di organi terminali che reagiscono al tatto e alla pressione sulla pelle. Il comune denominatore dei recettori di questo gruppo è la capacità di deformarsi sotto tensione, e si sa che molti di essi sono provvisti di congegni strutturali a forma di capsule che ne influenzano la sensibilità meccanica, ma in nessuno di tali casi sono conosciuti i processi molecolari a livello delle terminazioni stesse. Un terzo grande gruppo è formato dai termocettori. Come si è ricordato sopra, nuovi tipi, quali gli elettrocettori, sono stati scoperti nei Pesci.
d) Potenziale generatore.
Nel secondo stadio della recezione l'evento iniziale specifico deve essere trasdotto in un processo capace di stimolare il nervo afferente. Dato che tutti gli impulsi nervosi sono fondamentalmente simili e consistono in brevi potenziali d'azione (v. fig. 1; v. elettrofisiologia; v. neurone e impulso nervoso) non occorre che il secondo stadio sia specifico. Si è osservato che questo processo è lo stesso in tutti i casi e si manifesta come una variazione di potenziale elettrico attraverso la membrana del neurone sensoriale, il potenziale generatore di Granit (v., 1947). Nella maggior parte degli organi di senso il nervo afferente ha origine dentro o sul recettore, il cui potenziale di membrana, innescato dal processo iniziale specifico, è variato nel senso di una depolarizzazione. Questo processo elettrico (v. fig. 1) produce una corrente abbastanza intensa da stimolare il nervo a scaricare impulsi. La successione degli eventi è quindi: processo specifico-corrente generatrice-scarica nella zona di eccitazione della fibra nervosa. La facoltà di generare un potenziale d'azione appartiene alla membrana nervosa: qualora questa abbia subito una sufficiente depolarizzazione, genera un potenziale d'azione, cioè l'impulso nervoso, come nella fig. 1. L'organo terminale stesso non è generatore in questo senso e non risponde con potenziali d'azione alla propria depolarizzazione. Questi meccanismi saranno illustrati più avanti in maggior dettaglio (v. cap. 2, È a).
Quando la fibra nervosa afferente termina con una sinapsi sul recettore, come è mostrato chiaramente nella fig. 33, la corrente che si genera innesca di norma un processo di trasmissione chimica per il proprio trasferimento.
Da quando gli sperimentatori sono riusciti a registrare direttamente dall'organo terminale, il termine ‛potenziale del recettore' viene spesso usato per descrivere lo stesso evento, in alternativa a ‛potenziale generatore' il quale, in senso stretto, si riferisce alla depolarizzazione che stimola il nervo afferente. I due termini sono sinonimi per organi terminali semplici, ma in organi di senso complessi come la retina, che ha due strati di neuroni fra il recettore e il nervo (il nervo ottico), è corretto distinguere il potenziale del recettore da quello delle cellule gangliari che generano la corrente che stimola il nervo ottico.
Fra gli organi terminali dei Mammiferi la retina è un'eccezione, anche perché i potenziali del recettore sono iperpolarizzazioni della membrana cellulare da uno stato fondamentale di depolarizzazione o più alta permeabilità. Lo stadio successivo è un'elaborazione neurale di un equilibrio bilanciato di variazioni di potenziale nel ‛piccolo cervello' di neuroni attaccati ai recettori. Il potenziale generatore nel neurone finale o terzo neurone, la cellula gangliare, riassume il risultato di questa elaborazione del messaggio nel ‛cervello' retinico e lo traduce in impulsi. Gli altri organi terminali dei Mammiferi, per quel che se ne sa, per lo più rispondono con depolarizzazioni che generano impulsi nella maniera descritta. Recenti ricerche (v. sotto, cap. 2, È d) suggeriscono la possibilità di iperpolarizzazioni anche nei recettori olfattivi. Non si può però dire nulla su ciò che si scoprirà quando i numerosi organi di senso degli Invertebrati saranno studiati esaurientemente con le attuali potenti microtecniche intracellulari. Cosi nella chiocciola Aplysia sono stati trovati chemocettori che rispondono agli odori in due maniere diverse. Arvanitaki e altri (v., 1967) registrarono intracellularmente da un totale di 246 di queste terminazioni, la grande maggioranza delle quali rispondeva con potenziali generatori depolarizzanti del tipo ordinario, mentre 58 recettori reagivano con una primaria iperpolarizzazione, la quale ha un effetto opposto o inibitore sulla generazione degli impulsi. Le iperpolarizzazioni differivano da quelle nei bastoncelli e coni retinici per essere associate a una diminuzione invece che a un aumento della resistenza della membrana. Dal punto di vista del trasporto delle informazioni, l'inibizione può essere utile solo nel caso di un'eccitazione preesistente che l'iperpolarizzazione può sopprimere oppure liberare come scarica off al suo ritorno alla linea di base. Per il momento s'ignorano i dettagli di questo meccanismo.
È chiaro che le informazioni contenute nel processo iniziale specifico devono essere riprodotte nel potenziale generatore: l'intensità della stimolazione si rifletterà perciò nell'ampiezza della risposta del potenziale generatore, la sua velocità di salita e la sua durata nella velocità di salita e nella durata del potenziale generatore. La relazione quantitativa fra il processo specifico e il suo potenziale generatore può essere lineare o esponenziale, differente nei diversi organi di senso, ma una volta che il messaggio è stato trasdotto in una corrente che genera una scarica di impulsi, le proprietà di questo meccanismo generatore determinano direttamente la frequenza di scarica. Questa relazione lineare fra la grandezza di un potenziale depolarizzante una membrana e la frequenza di scarica della sua fibra nervosa è stata verificata sia per i neuroni centrali sia per gli organi di senso, e dimostrata generalmente valida. Fu studiata per la prima volta con un recettore di stiramento, un fuso neuromuscolare, nel muscolo del dito di rana (v. Katz, 1950): la registrazione era indiretta, in quanto il potenziale generatore era intercettato nella terminazione nervosa a una certa distanza dalla sua origine, cioè dopo una certa sua attenuazione causata dal trasporto elettrotonico fino a quel punto; tuttavia, fu possibile studiare la relazione fra il potenziale generatore e la frequenza degli impulsi e si dimostrò che essa segue la curva lineare della fig. 2 (v. cap. 2, È b).
Nella fig. 3 è illustrato il risultato di esperimenti, condotti mediante registrazione intracellulare da un certo numero di motoneuroni (v. Granit e altri, 1966), sulla relazione fra potenziali generatori, qui detti potenziali postsinaptici (v. Eccles, 1957), e frequenze di scarica dei neuroni: anche in questo caso la relazione è lineare. Per ottenere questa curva fu necessario fornire una stimolazione riflessa costante a un motoneurone prima e dopo che questo fosse eccitato dalla corrente transmembranale erogata dalla punta dell'elettrodo intracellulare: prima, per ottenere il valore del potenziale postsinaptico prodotto da uno stimolo riflesso in un neurone silente; dopo, per misurare di quanto lo stimolo riflesso aumentasse la frequenza degli impulsi del motoneurone eccitato rispetto alla frequenza di scarica fondamentale che era stata generata dalla corrente dall'interno della cellula attraverso la sua membrana.
Ovviamente, i potenziali postsinaptici misurati negli esperimenti della fig. 3 erano per lo più prodotti da stimolazione dei recettori; sorge il problema circa il modo con cui essi sono in rapporto con la frequenza degli impulsi che li hanno prodotti. In alcuni casi in cui è stato possibile studiare questo rapporto, si è osservato che gli impulsi che arrivano a un neurone producono potenziali generatori postsinaptici proporzionali alla loro frequenza e al loro numero totale.
Tutte queste semplici relazioni lineari sono state riscontrate valide entro un considerevole intervallo di normali frequenze di scarica delle terminazioni sensoriali e dei neuroni del midollo spinale. Esse dimostrano che, fondamentalmente, un'immagine dell'originale frequenza di scarica dell'organo terminale può essere trasmessa senza distorsioni a neuroni centrali nel cervello, anche se frequenza d'impulsi e potenziale generatore si sono alternati più volte nelle transizioni da un neurone all'altro lungo la via ascendente. Si conoscono trasformazioni lineari di questo tipo per i meccanocettori della pelle (v. Mountcastle, 1965) e per i fusi neuromuscolari (v. Jansen e Wallöe, 1970). D'altra parte, i messaggi da organi di senso complessi, come l'occhio e l'orecchio, sono ‛revisionati' e trasformati lungo le varie stazioni dall'interazione fra eccitazione e inibizione. Questo avviene in un modo che esalta i segnali utili nella discriminazione percettiva a scapito della meno importante, semplice funzione intensità-frequenza degli impulsi. La fedeltà con cui l'originale potenziale generatore sensoriale è riprodotto nelle stazioni superiori sarà alla fine leggermente modificata dal ‛guadagno' nelle membrane delle cellule cui è trasmesso. Per guadagno s'intende la costante di proporzionalità o la pendenza della curva che descrive il potenziale generatore in funzione della frequenza di scarica degli impulsi da esso generati: in altre parole il valore che nella fig. 3 è 2,28 impulsi/s/mV. Quindi, per es., in cellule più piccole di quelle dalle quali è stato registrato intracellularmente in quella particolare ricerca, il guadagno sarebbe stato minore (v. Kernell, 1966).
e) Adattamento.
Supponendo che la stimolazione sia costante e di lunga durata, sorgono i problemi del mantenimento e della stabilizzazione della scarica, riguardo sia ai potenziali generatori che alle frequenze dei potenziali di azione. I vecchi psicofisici del secolo scorso sapevano bene che gli organi di senso (o qualche stazione superiore) erano soggetti all'adattamento. Quando un pelo viene piegato e mantenuto nella nuova posizione, scompare quasi immediatamente la breve esperienza sensitiva di tatto. Una delle prime conquiste nell'era elettronica della ricerca sensoriale fu la localizzazione di un processo dominante di adattamento nell'organo terminale stesso, come è mostrato dalla schematica fig. 4 presa dai primi lavori di Adrian. Le due terminazioni sensitive più in alto nella figura (fuso neuromuscolare, pressione) si adattano lentamente e perciò sono in grado di dare risposte abbastanza ben mantenute; la coppia successiva (tatto, pelo) risponde fasicamente. Si supponeva che la fibra nervosa inclusa da Adrian nel diagramma fosse stimolata elettricamente. È stato sottolineato più sopra che il potenziale generatore di una terminazione sensoriale mantenuto per qualche tempo è senz'altro capace di mantenere stimolata la corrispondente fibra nervosa afferente. Una conseguenza importante dell'adattamento è che la maggior parte degli organi terminali è più sensibile alle variazioni e alla velocità di variazione dello stimolo adeguato che alla sua durata.
Il processo di adattamento potrebbe essere una caratteristica dell'evento iniziale specifico o del successivo potenziale generatore o di tutti e due. Il caso meno comune è forse l'adattamento nel processo iniziale. Tuttavia, un esempio di questo è la scissione, sotto l'influenza di una luce abbastanza forte, del fotopigmento retinico, la rodopsina (o porpora visiva): questa sostanza si trova nei bastoncelli retinici e quando è stata scissa sono i coni a funzionare da recettori visivi. Tale cambiamento, noto come adattamento alla luce, è un'esperienza quotidiana che si ha al mattino, al momento del risveglio. Anche eventi nervosi complessi contribuiscono all'adattamento alla luce. La rodopsina si rigenera al buio la sera, e i bastoncelli rientrano in scena come organi dominanti della visione crepuscolare. La rigenerazione della rodopsina è nota sotto il nome improprio di adattamento al buio.
In molti meccanocettori il processo di adattamento è inerente al potenziale generatore, senza però essere necessariamente una proprietà della membrana cellulare responsabile di questa variazione di potenziale. Più sotto (v. cap. 2, È a), si vedrà che nel corpuscolo del Pacini, che è una terminazione incapsulata, il processo di adattamento è dovuto quasi completamente all'incapacità della capsula di sostenere una stimolazione prolungata. Analogamente, nei fusi neuromuscolari del gatto v'è un tipo di organo terminale (v. cap. 2, È b), chiamato terminazione primaria, che è molto sensibile alla velocità con cui il muscolo è stirato, ma si adatta rapidamente a una frequenza di scarica molto più bassa durante una trazione continua. In questo caso l'adattamento è in gran parte dovuto a un mutuo gioco di forze elastiche e viscose nella fibra muscolare modificata (intrafusale), la cui porzione centrale contiene la terminazione stessa del fuso. Di per sé la membrana cellulare di quest'organo terminale nei Mammiferi sarebbe perfettamente in grado di sostenere una stimolazione prolungata. Tuttavia, una lenta caduta del potenziale di membrana è inevitabile nella maggior parte delle cellule sotto stimolazione continua. Ciò provoca una riduzione graduale della frequenza di scarica, meglio conosciuta da esperimenti eseguiti sui motoneuroni. La maggioranza delle terminazioni sensitive si è dimostrata meno accessibile delle cellule motorie del midollo spinale alle registrazioni intracellulari richieste per tali studi. Quindi spesso s'ignora dove ha luogo il processo principale di adattamento di una terminazione, e i fisiologi si sono dovuti accontentare di misurare il processo di adattamento in termini di una diminuzione della frequenza di scarica del recettore. Nella maggior parte dei meccanocettori l'adattamento probabilmente dipende in larga misura dal tessuto circostante, le cui proprietà - come negli esempi succitati - sono spesso integrate nella risposta totale della struttura reattiva.
f) Altri modi di trasmettere informazioni sensoriali.
Si dovrebbe accennare brevemente al fatto che la trasduzione di una risposta sensoriale in un potenziale generatore che produce impulsi nel suo nervo afferente non è l'unico modo di trasmettere il messaggio sensoriale. In un organo complesso come la retina, dove molte cellule hanno costruito un ‛cervello periferico' per l'immediata modulazione del messaggio, le cellule adiacenti si influenzano l'una con l'altra per interazione fra le variazioni positive o negative della loro corrente di membrana. È dubbio se in questo centro la produzione di impulsi abbia importanza, sebbene alcuni lavori riportino che si possono a volte osservare brevi ma intense scariche di impulsi quando il microelettrodo si trova su un neurone chiamato cellula amacrina (per una trattazione completa dell'argomento rimandiamo all'articolo visione); si accenna qui all'organizzazione retinica solo per introdurre questo particolare problema e per sottolineare che la produzione di impulsi e la conseguente perdita parziale di informazioni sono un processo necessario unicamente per la conduzione a lunga distanza dei segnali. All'interno della retina non esiste quasi questa necessità e perciò solo pochi impulsi sono generati prima del suo ultimo neurone, la cellula gangliare, i cui assoni formano il nervo ottico.
g) Campi recettivi.
La grande maggioranza delle fibre nervose afferenti possiede terminazioni su diversi organi terminali della stessa modalità. Registrare impulsi da una singola fibra nervosa afferente significa pertanto che il segnale proviene da un ‛campo recettivo' di terminazioni simili. Questo concetto è bene illustrato da uno dei primi risultati di Adrian ottenuto registrando da singole fibre afferenti del tatto nella pelle di rana (v. fig. 5). Nella figura si vede anche che campi recettivi di fibre differenti tendono a sovrapporsi nella distribuzione.
I campi recettivi sono da allora diventati uno degli argomenti preferiti di studio. È noto che essi variano molto anche in grandezza, da quelli puntiformi della pressione, le cui dimensioni sono comprese fra 1 e 2 mm2, a quelli estremamente ampi, fino a 30×90 mm, ottenuti con certe fibre afferenti rispondenti a leggero contatto (v. cap. 2, È c). Persino in un organo incapsulato come il fuso neuromuscolare, che contiene diverse terminazioni sensitive entro la stessa capsula, una singola fibra afferente ne innerva molte ma non raggiunge altri fusi dello stesso muscolo. La sovrapposizione di campi recettivi nella maniera e nella scala della fig. 5 sembra essere una caratteristica dell'innervazione della pelle.
Il concetto di ‛campo recettivo' assume un altro significato quando si riferisce alla retina. In questo organo i recettori convergono verso le cellule bipolari e queste ultime verso le cellule gangliari: una fibra del nervo ottico che si diparte da una cellula gangliare, quindi, registra l'effetto finale della convergenza totale modificato in altri tipi di cellule che distribuiscono lateralmente eccitazione o inibizione. Il campo recettivo di questa complessa organizzazione si estende in superficie come un campo recettivo cutaneo, ma è chiaramente un tipo molto differente di unità anatomica e fisiologica. Lo studio dei campi recettivi della retina s'è rivelato particolarmente importante nella moderna fisiologia della visione (v. visione).
Il termine ‛campo recettivo' è usato nello stesso senso che per la retina nelle ricerche sulle singole fibre o cellule d'un centro nervoso, qualora esse siano attivate da molte terminazioni della stessa modalità. Come avviene per le cellule gangliari della retina (o le loro fibre), i messaggi registrati da neuroni corticali sono spesso molto rielaborati o resi più netti nelle stazioni intermedie per maggiore utilità dell'organismo. Per esempio, larghe bande di colore o di tono nel recettore, modulate dall'interazione, spesso appaiono come stretti picchi che favoriscono la discriminazione di queste qualità.
h) Importanza della dimensione della fibra.
Non è raro che differenti modalità sensitive siano rappresentate in fibre afferenti riunite in gruppi di un particolare calibro in uno spettro di distribuzione dei diametri delle fibre dei nervi di cui fanno parte. Le fibre afferenti spinali, che secondo la suddivisione effettuata da Lloyd (v., 1943; v. tab. I) rientrano nei gruppi I-III, sono provviste di una guaina mielinica.

Erlanger e Gasser (v., 1937) avevano dimostrato che la velocità di conduzione del potenziale d'azione è proporzionale al diametro della fibra. Per ottenere la velocità di conduzione in metri per secondo, le cifre della tab. I si devono moltiplicare per 6. Molto lavoro è stato da allora dedicato alla misurazione degli spettri dei calibri di vari nervi afferenti; nei nervi misti tali misurazioni sono state effettuate dopo degenerazione della porzione afferente. L'animale da esperimento preferito è stato il gatto, i cui nervi muscolari e cutanei sono stati anche recentemente sottoposti a un'analisi molto dettagliata (v. Boyd e Davey, 1968). V'erano notevoli differenze fra singoli nervi muscolari, ma in genere le fibre del gruppo I avevano un diametro totale da 9 a 16 μm (con una media di 12,5 μm), le fibre del gruppo II da 3 a 10 μm (con una media di 6,5 μm). V'era perciò una certa sovrapposizione fra le fibre del gruppo I e del gruppo II nell'intervallo 9-10 μm. Le fibre del gruppo III avevano un diametro variabile da 1 a 5 μm (con una media di 3,0 μm). Negli istogrammi le zone di sovrapposizione erano contrassegnate da evidenti minimi.
Le fibre dei nervi cutanei mielinici appartenevano a due soli gruppi, uno da 4 a 14 μm di diametro, con una media di 9 μm, l'altro da 1 a 7 μm con una media di 4 μm, con curve di distribuzione tendenti a essere bimodali.
La rilevanza di queste cifre per l'argomento che trattiamo è imperniata sulla loro equivalenza funzionale, la velocità di conduzione, sull'uso che se ne può fare nel lavoro sperimentale per identificare i recettori, e sul significato che nei riguardi della trasmissione del messaggio alla stazione centrale riveste il fatto che gli impulsi arrivino celermente o siano leggermente ritardati. La complessa struttura dell'innervazione cutanea verrà presa in esame più avanti (v. cap. 2, È c). Per quanto riguarda le fibre afferenti muscolari, è generalmente possibile qualche differenziazione sulla base della velocità di conduzione. Così le afferenze primarie fusali si trovano sempre fra le fibre più rapide, le secondarie nel gruppo II. Gli organi tendinei del Golgi di molti muscoli sono veloci quanto le afferenze primarie fusali, in altri muscoli sono un po' più lenti (v. Granit, 1970). Si è osservato che le fibre afferenti muscolari del gruppo III, la maggior parte delle quali non trasporta impulsi di stiramento, registrano la pressione e il dolore e alzano la pressione sanguigna. Nelle fibre amieliniche del gruppo IV, che sono state studiate meglio nelle afferenze cutanee (v. cap. 2, È c), il problema dell'identificazione del recettore in base alle proprietà della fibra s'è rivelato molto complesso. Circa il significato della velocità di conduzione per l'elaborazione centrale del messaggio sensitivo, si sa poco di preciso: sembra però ovvio che quando messaggi dello stesso generico tipo sono portati sia da fibre rapide che da fibre lente, le prime preparino in qualche modo i centri per gli effetti di queste ultime.
i) Soglie.
La sensibilità degli organi terminali a stimoli adeguati è notevolmente elevata. Cosi, per esempio, nei Mammiferi i recettori di stiramento dei muscoli (fusi) rispondono ad allungamenti dell'ordine di 4-10 μm; le soglie termiche sono dell'ordine di 0,2 °C; le sostanze di attrazione sessuale di alcuni insetti provocano una risposta olfattiva comportamentale in concentrazioni di 10-12 μg per ml; i bastoncelli retinici sono attivati da un singolo fotone di luce e i meccanocettori dell'organo di Corti dell'orecchio rispondono a vibrazioni dell'ordine di una frazione del diametro dell'atomo di idrogeno. La pelle è sensibile a pesi di 2-3 mg, a seconda dell'area stimolata.
È difficile immaginare come sarebbe il mondo per un organismo vivente se a tutte le sue terminazioni sensoriali fosse sempre permesso di emettere impulsi in proporzione a queste enormi sensibilità. Questo tuttavia è impedito da diverse misure di sicurezza, molte delle quali sono facoltative nel senso che possono essere adottate o eliminate dall'organismo per il migliore funzionamento dell'insieme. Ben noto è un fattore centrale come l'‛attenzione', ma vi sono inoltre meccanismi automatici di modulazione pre-e postsinaptici a tutti i livelli dal midollo spinale in su. Tali meccanismi centrali esulano dal tema dei recettori veri e propri, tranne nei casi in cui agiscono perifericamente, sulle terminazioni sensoriali. Si conosce un controllo periferico attuato da speciali fibre nervose (fibre centrifughe) per i fusi neuromuscolari, la coclea, l'apparato vestibolare e la retina; alcuni esempi saranno presi in considerazione più avanti (v., per es., cap. 2, È b).
2. Recettori specifici.
a) Il corpuscolo di Pacini.
Il problema fondamentale dei potenziali generatori in rapporto alla generazione di impulsi e all'adattamento è stato studiato in modo esemplare, in questo organo terminale, da W. R. Loewenstein. Una delle ragioni di successo in questo campo è rappresentata dalle insolite dimensioni di questo recettore, un corpo ellissoidale di 1,0×0,6 mm, che offre possibilità uniche per lo studio. Questo corpuscolo è un puro meccanocettore formato da lamelle (membrane) concentriche che circondano una terminazione amielinica d'una fibra nervosa afferente, per il resto mielinica (v. fig. 6). In effetti la mielinizzazione del nervo comincia subito dentro la capsula dove si trova il primo nodo di Ranvier. I corpuscoli di Pacini sono presenti nei tendini, nelle fasce muscolari e nella pelle, ma sono più facilmente individuabili nel mesentere (per es. del gatto), nel quale si possono vedere a occhio nudo come corpi ovoidali leggermente opachi in un mezzo trasparente.
La loro sensibilità altamente specifica alla pressione è dell'ordine di 0,2-0,4 μm, come si può calcolare mediante l'appena visibile potenziale generatore.
La loro risposta è fasica. Precedenti lavori (v. Gammon e Bronk, 1935) avevano già stabilito che i corpuscoli di Pacini del mesentere rispondono con impulsi a piccolissime pulsazioni dei vasi mesenterici. Loewenstein e i suoi collaboratori (v. Loewenstein, 1958; v. Loewenstein e Mendelson, 1965; v. Loewenstein e Skalak, 1966) hanno messo a punto una tecnica per studiare quest'organo in microscopia a contrasto di fase: sono riusciti, scortecciando lamelle concentriche, a eliminare virtualmente il 99,9% del volume del corpuscolo, fino a farne rimanere solo un nucleo centrale di 25 μm di diametro, contenente la terminazione nervosa circondata da alcune lamelle non più asportabili; premendo sul nucleo così isolato, si riusciva a ottenere un potenziale generatore normale (v. fig. 7A).
Se si seziona il nervo e si lasciano passare alcuni giorni perché si manifesti la degenerazione del moncone periferico, la risposta di generatore (identificabile in questo organo col potenziale del recettore) scompare, il che prova che la struttura sensibile è la terminazione nervosa stessa. Persino piccoli frammenti del nucleo di un normale corpuscolo di Pacini rispondono con potenziali generatori. Tale stimolazione frazionata della terminazione decapsulata fu resa possibile da una puntina attaccata a un cristallo piezoelettrico: facendo funzionare il cristallo con un campo elettrico si ottenne una sfera di compressione di 45 μm che era proporzionale all'intensità del campo. Mentre questo strumento serviva da trasduttore di energia elettrica in meccanica, l'organo terminale stesso trasduceva l'energia meccanica in elettrica. La corrente elettrica fornita dal generatore stimola la fibra nervosa in corrispondenza del primo nodo di Ranvier dove, nell'attraversare la membrana verso l'esterno, produce la scarica di impulsi.
Il ruolo di questa capsula lamellata come una cipolla è quello di costituire uno strumento per l'applicazione della pressione. Le lamelle solidamente attaccate al nucleo sono circondate da quelle asportabili, provviste di interstizi che aumentano a seconda del diametro conferendo così alla capsula la sua forma arrotondata e la sua grandezza. Gli interstizi sono riempiti da un liquido viscoso, ma poiché le lamelle sono connesse fra loro da elementi che cedono elasticamente, lo spostamento statico di una lamella influenza anche quelle situate a una certa distanza. In assenza di questo complesso sistema di lamelle, l'organo terminale risponde alla pressione come nella fig. 7A, mentre il corpuscolo normale intatto produce un potenziale generatore che sale in un paio di millisecondi e cade a zero con una costante di tempo dell'ordine di 3-5 ms. Alla cessazione della compressione l'organo produce un secondo potenziale generatore che è una risposta di cessazione (off; v. fig. 7B).
Così nel corpuscolo di Pacini intatto la maggior parte del rapido adattamento a uno stimolo continuo, che si riflette nel potenziale generatore, è causata dalle proprietà filtranti della capsula. Ugualmente, la sorprendente risposta di cessazione (off) dipende dall'esistenza di una capsula, e sembra dovuta alla liberazione dell'energia immagazzinata nelle lamelle elastiche. Pertanto si può dire che il ruolo della capsula è quello di modificare il potenziale generatore in modo che la risposta statica dello stelo terminale venga sostituita da una risposta fasica on-off. Questo rende il corpuscolo di Pacini uno strumento sensibile per la registrazione delle vibrazioni ad alta frequenza. Il suo rapido adattamento allo stimolo non ha nulla a che vedere con le proprietà del recettore; è una proprietà del tessuto capsulare circostante, e lo stesso si può dire di molti altri meccanocettori nelle articolazioni e nella pelle, studiati meno a fondo di questo. Un certo grado di adattamento molto lento alla produzione di impulsi da parte della membrana nervosa generatrice esiste, come ricordato in precedenza, in tutti gli organi. (Per una completa trattazione teorica dell'effetto capsulare, si rimanda al lavoro di Loewenstein e Skalak, 1966).
b) Il fuso neuromuscolare dei Mammiferi.
Quest'organo terminale incapsulato servirà nel presente contesto come esempio di una struttura sotto controllo centrifugo di speciali fibre nervose per mezzo delle quali la sensibilità dell'organo può essere modificata in rapporto alle necessità della locomozione e della postura. Esso è un recettore di stiramento che misura la lunghezza e la velocità di variazione della lunghezza del proprio muscolo. Un gran numero di fusi neuromuscolari si trova nei muscoli la cui lunghezza deve essere ben controllata, pur non subendo grandi variazioni (muscoli delle dita e del collo). Ma fusi neuromuscolari sono presenti virtualmente in tutti i muscoli striati.
Lo schema della fig. 8 mostra un fuso neuromuscolare con la propria fibra nervosa afferente avvolta intorno a un sacco elastico che contiene i nuclei (sacco nucleare). In A il fuso scarica impulsi, perché v'è una certa tensione nel sacco che è sospeso fra i due poli di una fibra di un muscolo striato, chiamata fibra intrafusale per distinguerla dall'ordinaria muscolatura extrafusale circostante. Quando le fibre extra- e intrafusali sono allungate contemporaneamente (v. fig. 8B), il fuso neuromuscolare scarica impulsi. Dopo (v. fig. 8C) occorre un certo tempo perché la fibra intrafusale si riporti alla lunghezza originale e all'originale frequenza di scarica. Se il muscolo extrafusale si contrae, il fuso viene scaricato (v. fig. 8D) e alla sua scarica segue una pausa. Ritornato alla lunghezza originale (v. fig. 8E), riprende lentamente la propria frequenza di scarica. Le proprietà di scarica descritte sono caratteristiche della cosiddetta afferenza primaria del fuso neuromuscolare. V'è un altro recettore, l'afferenza secondaria, con bassa sensibilità alla velocità di variazione di lunghezza, ma con una sensibilità molto simile a quella dell'afferenza primaria alla distensione statica. La descrizione che segue sarà dedicata principalmente alla terminazione primaria. Entrambi gli organi terminali e la loro innervazione afferente sono mostrati nella fig. 9, che illustra inoltre l'esistenza di due tipi di fibre intrafusali, chiamate rispettivamente fibra a sacco nucleare e fibra a catena di nuclei. I nomi derivano dal modo in cui i nuclei sono organizzati nei due tipi di fibre intrafusali.
In ogni fuso neuromuscolare v'è generalmente un certo numero di fibre intrafusali a sacco nucleare e a catena di nuclei, che sono tutte tenute insieme da una capsula a forma di fuso: a questa si deve il nome che è stato dato all'organo. La terminazione caratteristica sulle fibre a sacco nucleare del fuso è quella primaria, ma il suo nervo afferente invia anche un ramo alla fibra a catena di nuclei (v. fig. 9). Questa sembra essere la disposizione regolare. Analogamente la terminazione secondaria registra la risposta sia delle fibre intrafusali a sacco che di quelle a catena, ma in questo caso l'effetto assolutamente dominante proviene dalla fibra a catena. Il ramo della terminazione secondaria alle fibre intrafusali a sacco sembra essere raro.
Il controllo centrifugo è esercitato dalle fibre motrici dirette ai muscoli intrafusali, principalmente dalle fibre gamma di Leksell (v., 1945), che spesso vengono anche designate come fibre fusimotrici. Queste sono di due tipi (v. Matthews, 1962), dinamiche e statiche, e il loro effetto si può studiare nella fig. 10. In A viene illustrata l'estensione meccanica del muscolo, in B il comportamento dell'afferenza primaria in assenza di stimolazione; si noti l'aumento iniziale della frequenza di scarica durante la fase dinamica e la frequenza di scarica più bassa durante la distensione statica. In C si mostra come una fibra gamma statica, stimolata sin dall'inizio della registrazione, aumenti immediatamente la frequenza di scarica. Successivamente l'aumento è proporzionale alla distensione statica del muscolo nell'istante considerato. La stimolazione della fibra gamma dinamica in D ha una piccola influenza sulla scarica degli impulsi nel momento della distensione iniziale, mentre l'effetto principale si ha durante la fase dinamica dell'allungamento. Vi sono più fibre statiche che dinamiche distribuite ai fusi neuromuscolari dei Mammiferi finora studiati. Sembra che i due tipi di fibre gamma diano al fuso la sensibilità alla posizione (fibre gamma statiche) e alla velocità (fibre gamma dinamiche). Inoltre alcuni fusi neuromuscolari sono innervati anche da collaterali di fibre motrici che innervano la muscolatura extrafusale.
Per ciò che riguarda le fibre intrafusali, pare che le fibre gamma statiche siano distribuite alle fibre intrafusali a catena e pertanto influenzino le afferenze primarie per mezzo dei loro rami che si distribuiscono a queste terminazioni intrafusali. Le fibre dinamiche vanno esclusivamente alle fibre intrafusali a sacco nucleare. Dato che le terminazioni secondarie sulle fibre intrafusali a sacco nucleare sono pochissime, se pure vi sono, l'azione dinamica è virtualmente ristretta alle fibre afferenti primarie. La loro elevata sensibilità alla velocità di variazione di lunghezza (velocità di stiramento) dipende in gran parte dalla componente elastica del sacco nucleare; la rapida caduta della frequenza di scarica, appena l'allungamento è completato (v. fig. 8C), è una conseguenza del lento adattamento della porzione muscolare viscosa della fibra intrafusale a sacco. Questo è un nuovo esempio di un fenomeno di adattamento dipendente in larga misura da fattori estranei al recettore stesso.
Tuttavia, è stata l'innervazione motoria del fuso neuromuscolare a sollevare recentemente tanto interesse verso questo organo, perché ha costretto a prendere in considerazione il fuso nella sua funzione di regolatore della postura e della locomozione piuttosto che come un organo produttore di informazioni sensitive soltanto. I risultati di alcuni esperimenti fanno pensare che le afferenze dei fusi neuromuscolari contribuiscano alla percezione della forza. La loro funzione meglio conosciuta, tuttavia, è in rapporto con le attività motrici, riflesse e volontarie. Per capire l'azione principale delle afferenze primarie fusali controllate dalle fibre gamma è necessario considerare alcuni atti motori in cui quest'organo è implicato.
Azioni con e senza controllo gamma possono essere esaminate nelle due registrazioni della fig. 11. Quella inferiore (C, D) mostra la risposta allo stiramento del muscolo gastrocnemio dopo recisione della sua innervazione motoria. La terminazione primaria passiva in questo muscolo subisce un lento adattamento dopo la fase dinamica ed è silente sia prima sia dopo l'atto dello stiramento. Le due registrazioni superiori (A, B) mostrano lo stesso fuso prima che le sue fibre gamma fossero recise per sezione dell'efferenza motoria dal midollo spinale. Vi è quindi attività continua a riposo, e un accurato studio ha dimostrato che nel gatto decerebrato il fuso era attivato sia da fibre gamma statiche che dinamiche. L'allungamento nello stiramento era lo stesso nei due casi, ma nell'animale non denervato il muscolo si contraeva in un riflesso di stiramento registrato dall'estensimetro cui era connesso. Questa è la causa della maggiore escursione del miografo (registratore di tensione).
Il riflesso di stiramento (v. Liddell e Sherrington, 1924) è trasmesso al midollo spinale dalle afferenze primarie fusali nel modo illustrato nella fig. 12, con una proiezione monosinaptica direttamente sulla cellula motoria (motoneurone) che determina la contrazione di fibre extrafusali del muscolo a cui appartiene il fuso: questa cellula viene chiamata generalmente motoneurone alfa. Come conseguenza lo stiramento del muscolo porta all'eccitazione dei suoi motoneuroni che causano una risposta di accorciamento del muscolo stesso per contrazione. Questa specie di accoppiamento è conosciuto come feedback negativo: negativo in quanto l'effetto retroattivo del riflesso sul muscolo, producendo accorciamento, si oppone all'allungamento. Il diagramma mostra altresì che lo sperimentatore ha la possibilità di verificare il comportamento della scarica fusale isolando una fibra nervosa nella radice dorsale, nel punto in cui essa penetra nel midollo spinale. L'estensimetro funziona da miografo per la registrazione del cambiamento riflesso di tensione. Nello schema è rappresentato anche il motoneurone gamma che determina la contrazione delle fibre muscolari intrafusali del fuso. Le linee che arrivano ai motoneuroni alfa e gamma dall'alto mostrano che entrambe le vie motorie possono essere attivate da varie strutture sopraspinali, ivi compresi i centri motori del cervello e del cervelletto.
Uno sguardo allo schema mostra che se il motoneurone gamma è attivato, e i poli muscolari del fuso sono contratti, il sacco nucleare verrà disteso proprio come lo era quando veniva stirato il muscolo nell'esperimento della fig. 11. È quindi possibile produrre un riflesso da stiramento anche in questa maniera indiretta attraverso il cosiddetto circuito gamma. Ovvero, mantenendo una scarica permanente nel motoneurone gamma, è possibile aumentare la sensibilità della terminazione sensoriale allo stiramento. Nella vita normale di un animale la maggior parte degli atti motori coordinati impegnano i motoneuroni alfa e gamma simultaneamente, in quella che è stata chiamata la coattivazione a opera del legame alfa-gamma. Il termine implica che in qualche parte del sistema nervoso centrale qualche tipo di legame fa funzionare insieme i due sistemi motori. Recentemente è diventato possibile registrare nell'uomo con microelettrodi dalle afferenze primarie fusali.
La fig. 13 mostra un esempio di legame alfa-gamma: durante il movimento volontario la scarica in un'afferenza primaria del muscolo che si contrae (riga superiore) avviene assieme alla contrazione extrafusale (terza riga), la cui risposta elettrica è registrata nella quarta riga. La seconda riga è una registrazione di un frequenzimetro che dà la frequenza di scarica istantanea del fuso. Tutte le registrazioni sono simultanee. Quindi, anche nell'attività volontaria v'è attivazione automatica dei motoneuroni gamma insieme con i motoneuroni alfa. Lo stesso vale per la maggior parte dei riflessi. Per queste ragioni lo studio dei fusi neuromuscolari ha modificato radicalmente le nostre idee su come il sistema nervoso centrale regoli il movimento dei muscoli nella locomozione e nella postura.
Diamo tre esempi della funzione delle afferenze primarie fusali nel controllo motorio: il primo riguarda il mantenimento di una contrazione posturale stabile. La lunghezza del muscolo sarà allora mantenuta fissando a un certo livello la sensibilità del fuso neuromuscolare. Se il muscolo si accorcia, si avrà diminuzione del carico applicato ai fusi, con conseguente diminuzione della loro attività. I motoneuroni extrafusali ricevono un numero ridotto di impulsi e il muscolo si allunga un poco. Però, dato che sono attivi sia i motoneuroni alfa sia i gamma, l'allungamento provoca un riflesso di stiramento dovuto alla trazione sul fuso che, dobbiamo ricordare, ha una sensibilità di soglia dell'ordine di 10 μm. Il muscolo è così obbligato a contrarsi, allentando di nuovo la trazione sui fusi, e in questo modo subirà piccole oscillazioni continue intorno alla lunghezza predisposta. L'organo di senso ha allora funzione analoga a quella di un termostato che, controllando una sorgente d'energia che genera calore, mantiene la temperatura a un valore costante: il fuso neuromuscolare è il sensore e il muscolo scheletrico è la sua sorgente d'energia. In entrambi i casi la stabilità è mantenuta da un'oscillazione intorno al valore medio.
Il secondo esempio riguarda ciò che è stato chiamato compensazione di carico. Supponiamo che un muscolo in contrazione incontri inaspettatamente una resistenza, ad esempio per un aumento del carico a cui è sottoposto. Nel movimento, che ha luogo sotto il controllo del legame alfa-gamma, i due circuiti (alfa e gamma) sono stati regolati per una precisa quantità di eccitazione. I fusi sono pertanto regolati in modo da controbilanciare con la loro contrazione la diminuzione di carico prodotta dall'aumento della contrazione extrafusale (accorciamento). L'aggiunta del carico inaspettato rende i fusi relativamente troppo attivi, in quanto essi erano regolati per compensare una diminuzione di carico che non si è verificata.
I fusi aumenteranno perciò la loro frequenza di scarica per resistere all'allungamento; se questo dovesse tuttavia verificarsi - a causa d'un carico elevato - determinerà uno stiramento del fuso e in tal modo lo ecciterà, provocando così una contrazione atta a controbilanciare la distensione. In tutti e due gli esempi, naturalmente, il riflesso di stiramento è lo strumento che esercita queste regolazioni automatiche.
Il terzo esempio illustra il ruolo della sensibilità alla velocità dell'organo terminale primario del fuso, controllato dalle fibre gamma dinamiche. Un riflesso di stiramento controllato esclusivamente dalla sensibilità alla posizione statica dovrà percorrere il circuito riflesso (v. fig. 12) prima che possa verificarsi un effetto sulla contrazione del muscolo. Questo significa che il contributo del fuso all'atto motorio sarà ritardato rispetto al segnale di agire a causa del tempo di conduzione lungo il circuito riflesso. La contrazione del muscolo e il segnale di contrazione del fuso saranno sfasati e ciò causerà oscillazioni disordinate del muscolo. La sensibilità alla velocità fa percorrere al segnale proveniente dal fuso il circuito riflesso mentre il muscolo è ancora nella sua fase ascendente di azione, e ciò impedisce una risposta oscillatoria. Il fatto che la sensibilità alla velocità sia controllabile può di conseguenza essere considerato come un mezzo di correzione del ritardo dovuto alla conduzione.
L'importanza delle fibre secondarie del fuso nel controllo motorio è attualmente meno conosciuta. Esse potrebbero essere responsabili principalmente della trasmissione ai centri superiori delle informazioni circa la lunghezza istantanea del muscolo che si contrae.
Nei muscoli v'è un terzo recettore di stiramento, conosciuto come organo tendineo del Golgi, che si trova nella zona di transizione fra muscolo e tendine ed è quindi in serie con le fibre del muscolo extrafusale che si contrae: esso è perciò uno strumento sensitivo per registrare la tensione di contrazione. Può essere interessante notare che l'organo tendineo ha sul motoneurone un effetto opposto a quello dell'afferenza primaria fusale: esso infatti produce inibizione là dove il fuso produce eccitazione, ed eccitazione dove il fuso produce inibizione, cioè sugli antagonisti al muscolo in cui il fuso stesso si trova. Il principio generale è tuttavia lo stesso per il fatto che anche l'organo tendineo agisce come un feedback negativo. Esso registra la tensione del proprio muscolo e ne produce il rilasciamento. Non v'è nessun sistema gamma per regolare la sensibilità degli organi tendinei. Se e in quale misura essi devono funzionare è determinato da circuiti posti interamente dentro il midollo spinale e controllati dall'alto.
(Per una trattazione degli organi terminali dei muscoli e dei loro meccanismi di controllo v. Granit, 1955, 1966 e 1970; v. Matthews, 1962 e 1964).
c) Recettori cutanei.
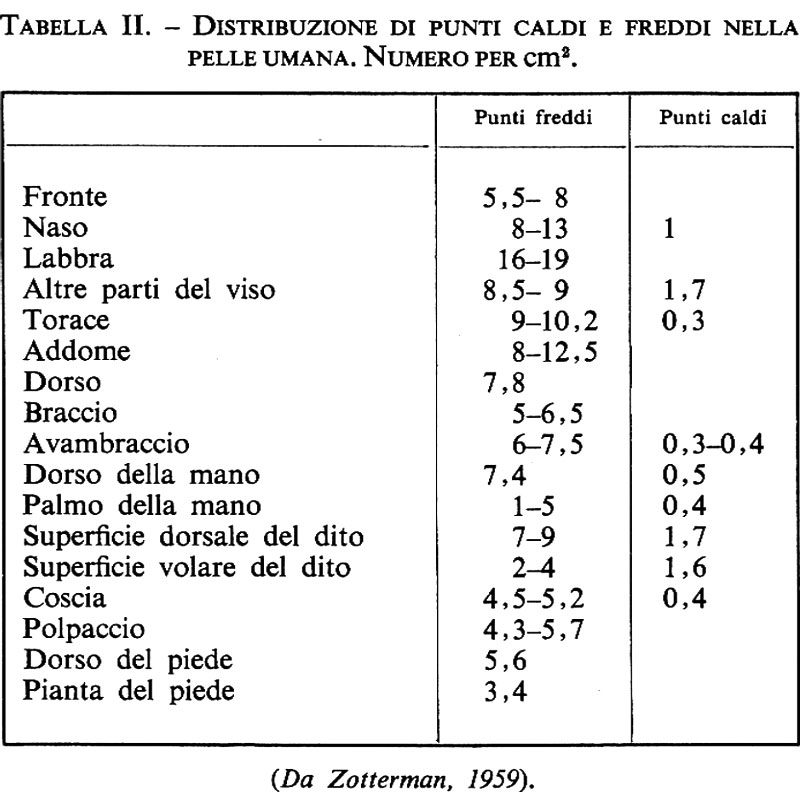
I contributi psicofisici ai problemi dei recettori della pelle cominciarono con la scoperta di Blix (v., 1882-1883) della rappresentazione puntiforme del freddo, del caldo e della pressione sulla pelle. Numerosi studi seguirono nella sua scia e può essere interessante riprodurre come esempio una tavola (v. tab. II), compilata da Zotterman (v., 1959), che mostra la distribuzione dei punti caldi e freddi nella pelle umana, mettendo in evidenza la maggior densità dei punti freddi rispetto a quelli caldi. Molte delle vecchie ricerche si occupavano delle correlazioni fra strutture della pelle, come si conoscevano allora, e sensazioni specifiche. L'era elettrofisiologica di ricerca ha affrontato lo stesso problema con la registrazione diretta; gli strumenti o ‛misuratori' sensitivi della pelle sono stati congegnati sulla base delle risposte specifiche al tatto, al caldo o al freddo. Non è necessariamente vero che si possano tradurre i loro messaggi, così come sono registrati direttamente, negli equivalenti sensitivi, così come sono percepiti. Più sotto, trattando dei recettori e quindi della periferia, potremo descrivere solo brevemente le correlazioni con le percezioni.
Il problema della specificità dei recettori cutanei rimase per qualche tempo confuso per il fatto che zone della pelle con e senza terminazioni incapsulate davano le stesse quattro risposte sensitive (v. Weddell e altri, 1954), il che indusse alcuni studiosi a pensare che la specificità non fosse che una questione di forme combinate di stimoli. Esperimenti recenti hanno unanimemente confermato la fondamentale validità del concetto classico della specificità dell'organo terminale a stimoli adeguati. Le nuove prove sono basate sulla registrazione da singole fibre afferenti, ora estesa persino alle piccole fibre C amieliniche a lenta conduzione del gruppo IV, rese accessibili dai recenti miglioramenti delle tecniche di registrazione (v. Douglas e Ritchie, 1957; v. Douglas e altri, 1960; v. Iggo, 1958). Molti organi terminali sono stati identificati precisamente. Avendo visto più sopra che la terminazione nervosa isolata del corpuscolo di Pacini risponde alla pressione e sapendo che le fibre afferenti sottili rispondono al caldo e quelle di maggiore spessore al freddo (v. von Euler, 1947), non v'è alcun ostacolo fondamentale all'attribuzione di sensibilità specifiche alle terminazioni nervose prive di strutture incapsulate. Nella pelle prevalgono le terminazioni libere di questo tipo. In realtà il lavoro di Zotterman (v.,1939), illustrato nella fig. 14, aveva mostrato già molto tempo fa che con i più semplici tipi di stimoli meccanici si ottenevano impulsi di velocità di conduzione molto differenti (ascissa). L'intera gamma delle velocità di conduzione delle fibre afferenti mieliniche è difatti rappresentata nella fig. 14.
Indipendentemente dal tipo di animale o dal tipo di pelle, sia pelosa sia glabra, i meccanocettori si dividono in due gruppi principali: quelli che si adattano lentamente e quelli che si adattano rapidamente. Conosciamo meglio e più in dettaglio quelli che si adattano lentamente, cioè quelli che Brown e Iggo (v., 1967) e Iggo e Muir (v., 1969) chiamano di tipo I. Di questo tipo è il corpuscolo illustrato nella fig. 15, elevato un po' sopra la superficie come una cupola ricoperta da un solo strato di cellule che differiscono dal resto delle cellule epidermiche. È conosciuto da molto tempo e descritto come le Tastzellen di Merkel. Si nota che il corpuscolo contiene cellule tattili provviste di una placca nervosa e innervate da una fibra mielinica relativamente grossa che può alimentare da uno a cinque corpuscoli di questo tipo. La velocità media di conduzione della fibra è, nel gatto, di 57 m/s. Quando si fa passare sul corpuscolo una sonda con un diametro alla punta di 0,25-2 mm, questo risponde fasicamente con frequenze di scarica che arrivano fino a 1.500 impulsi al secondo. La frequenza di impulsi di questa fase dinamica dipende dalla velocità e dall'ampiezza dello spostamento. Questa fase è seguita da una fase statica di scarica a frequenze più basse, che dura fino a 30 minuti o più se lo spostamento è sufficiente. Questo organo terminale è silente in assenza di stimolazione. La sua sensibilità è uniforme nell'area centrale, ma diminuisce verso il bordo della cupola e scompare nella pelle intorno al corpuscolo.
La seconda classe di meccanocettori a lento adattamento, tipo II di Iggo, ha una scarica regolare a riposo e una componente dinamica molto minore del tipo I. Le sue frequenze di picco sono più basse per spostamenti uguali delle due terminazioni. Lo stiramento laterale della pelle altera la sua sensibilità ma non ha alcuna influenza sulla scarica della terminazione del tipo I. La terminazione del tipo II è ben definita da queste proprietà, ma non è stata ancora identificata strutturalmente. La velocità di conduzione delle sue fibre mieliniche è un po' inferiore a quella del tipo I.
Entrambi questi recettori a lento adattamento differiscono da quelli a rapido adattamento per il fatto che sono sensibili anche alla stimolazione da freddo, il tipo II più del tipo I, ma l'azione della temperatura è debole in paragone con la potenza della stimolazione meccanica e non si ritiene rappresentativa di alcuna reale specificità bimodale o doppia.
I recettori dei peli appartengono tutti al gruppo dei recettori a rapido adattamento. Le loro proprietà variano a seconda del tipo di pelo. I peli più corti, fini e ondulati, sono i più numerosi: spuntano da un comune orifizio in gruppi di due o tre e hanno la più elevata sensibilità al movimento, rispondendo a un lieve soffio o allo strofinio di un batuffolo di cotone sulla pelliccia (nel gatto). I loro campi recettivi variano da 0,5 a circa 6 cm2 e le velocità di conduzione delle fibre afferenti sono le più basse (v. fig. 16), appartenendo al cosiddetto gruppo delta delle fibre mieliniche A (v. Erlanger e Gasser, 1937). I peli più lunghi, più grossi, dritti o leggermente curvi (guard hairs: monotrichi, peluria superiore), che spuntano singolarmente dai loro follicoli, scaricano 3-4 impulsi per unità, a qualsiasi velocità di spostamento; quando però si strofina la pelliccia con una bacchetta di vetro, si ottengono brevi e intense scariche di 600 impulsi al secondo. I loro campi recettivi vanno da 0,5 a 6,0 cm2 e ogni follicolo è innervato da numerose fibre afferenti che si sovrappongono in modo che peli G adiacenti possono essere componenti di unità differenti. Le loro velocità di conduzione sono mostrate nella fig. 16. I peli G del gatto sono stati recentemente suddivisi in due gruppi (v. Burgess e altri, 1968). I tylotrichs (peli T) sono i più lunghi e grossi di tutti: ognuno di essi emerge da un follicolo circondato da una rete di vasi sanguigni capillari, e ciascuna unità afferente alimenta da 3 a 10 follicoli T. Al movimento per stimolazione meccanica questi peli scaricano una serie di impulsi. L'unità del follicolo T non è molto comune nel gatto. Il suo campo recettivo tende a essere più vasto di quello degli altri due e la sua velocità di conduzione (v. fig. 16) è la più elevata fra i recettori dei peli. L'innervazione dei peli è illustrata nelle figg. 17 e 18.
Un quarto tipo di recettore cutaneo rapido è il corpuscolo di Pacini, le cui proprietà, fra le quali la sensibilità alla vibrazione, sono state descritte più sopra. Strutture meno definite sono i ‛recettori di campo'. È probabile che siano non omogenei, perché la velocità di conduzione afferente varia notevolmente, da 40 a 70 m/s; i più veloci manifestano un'attività sostenuta e rispondono alla vibrazione di 540 cicli/s, ma sono silenti a riposo; quelli le cui fibre hanno velocità di conduzione più lenta, più difficilmente mostrano sensibilità alla vibrazione e attività sostenuta. Hanno campi recettivi ovali e piuttosto ampi.
Quando si sono potute applicare le nuove microtecniche alle singole fibre amieliniche C, ci si è resi inaspettatamente conto che le informazioni provenienti da meccanocettori altamente sensibili sono trasportate in un gran numero di tali fibre. Solo recentemente è stato accertato che le fibre amieliniche costituiscono l'80% di tutti i nervi cutanei. In una recente ricerca su 131 fibre C, Bessou e Peri (v., 1969) individuarono 47 meccanocettori a bassa soglia (36%), in grado di essere eccitati già da una stimolazione puntiforme della pelle (gatto) con forze al di sotto di 1,0 g. Si è osservato anche che essi scaricano per qualche tempo dopo la stimolazione (v. Hensel e altri, 1960). La velocità di conduzione delle fibre C è al di sotto di 2 m/s.
I recettori cutanei vengono ora studiati nell'uomo introducendo microelettrodi direttamente nel nervo intatto (v. Hagbarth e Vallbo, 1969), e questo metodo ha reso possibile correlare le risposte bioelettriche alle esperienze sensitive prodotte dagli stessi stimoli. Questa nuova tecnica, detta microneurografia, è stata ben descritta da Hagbarth (v., 1976). Un altro importante progresso tecnico riguarda il controllo dell'ampiezza e della rapidità di aumento dell'incavatura della cute, ottenuta con tecniche adatte di stimolazione, allo scopo di mettere in rapporto lo stimolo meccanico con la frequenza di scarica delle singole fibre che rispondono allo stimolo pressorio. Questa tecnica utilizza un controllo a feedback dei parametri meccanici della stimolazione. Essa è stata utilizzata ampiamente da Valibo e collaboratori per lo studio dei meccanocettori della mano dell'uomo (cute glabra). Un recente simposio (v. Zotterman, 1976) tratta di questo argomento. La sensibilità tattile della cute provvista di peli è mediata da fibre mieliniche spesse, dotate di alta velocità di conduzione (v. Hallin e Torebjörk, 1976).
Quattro tipi di unità, due a rapido e due a lento adattamento, sono stati trovati nella mano dell'uomo da Burgess e Pearl, con risultati essenzialmente simili a quelli ottenuti nei Mammiferi al di sotto dell'uomo. Dei due tipi di meccanocettori a rapido adattamento, uno ha le proprietà del corpuscolo di Pacini, l'altro può essere probabilmente identificato col corpuscolo di Meissner, descritto da parecchi autori nella cute glabra della scimmia. Di notevole interesse è l'osservazione (v. Knibestöl e Vallbo, 1970) che su 60 singole fibre provenienti dalla mano dell'uomo, circa il 75% appartiene al tipo a lento adattamento, mentre nella scimmia circa la stessa percentuale è formata da unità a rapido adattamento. Vallbo (v., 1970) pensa che questo sia uno sviluppo che facilita l'esplorazione tattile da parte della mano dell'uomo. I campi recettivi variano come è mostrato nella fig. 19.
Poiché le unità a rapido adattamento e quelle a lento adattamento del tipo I hanno campi recettivi piccoli e ben delimitati, distribuiti in modo da permettere interdigitazione con profili di sensibilità ben conservati, esse appaiono manifestamente adatte alla discriminazione spaziale e alla localizzazione accurata degli stimoli. I corpuscoli di Pacini e le unità del tipo II a lento adattamento mancano di queste proprietà, e pertanto si ritiene che esse rispondano alle vibrazioni meccaniche che passano sopra la cute e al suo stiramento. Ma per queste correlazioni psicofisiche, in rapporto a stimoli soprasogliari, occorre anche considerare le risposte di popolazioni piuttosto che quelle di singole unità. Anche piccole variazioni dell'intensità o della localizzazione dello stimolo determinano apprezzabili cambiamenti nei tipi di risposta, dovuti al coinvolgimento di terminazioni adiacenti.
Quanto al dolore, si sa da tempo (Bishop, Casser, Ranson, Zotterman) che stimoli che per analogia interpretiamo come dolorosi, in quanto provocano innalzamento della pressione sanguigna, contorcimenti e vocalizzazione negli animali, sono condotti dalle fibre mieliniche di diametro più piccolo e dalle fibre C (v. de Reuck e Knight, 1966). Nei lavori più recenti sulle singole fibre afferenti isolate con microelettrodi l'indice di dolore è stato la presenza di impulsi in fibre ad alte soglie meccaniche, per stimoli quali la puntura d'ago, il pizzicotto della pelle, ecc.; il tatto o la pressione da soli non sono riusciti a eccitarli. Di 147 fibre afferenti cutanee amieliniche, circa un terzo ha risposto soltanto o più intensamente a stimoli meccanici intensi con carattere nocivo (v. Bessou e Perl, 1969). Non v'è oggi alcuna prova in favore del vecchio assunto che il dolore sia puramente una risposta a una stimolazione eccessiva di un qualsiasi tipo di recettore cutaneo. Tutta la ricerca più recente sostiene il concetto fondamentale di specificità delle terminazioni nervose, anche per il tipo di stimolo che può chiamarsi nocivo, per quanto sia stata descritta un'eccitabilità polimodale di qualche terminazione.
Una recente teoria, secondo la quale il dolore nascerebbe da processi d'interazione al livello dell'entrata nel midollo spinale delle fibre afferenti, non è stata accettata da nessuno fra i maggiori sperimentatori in questo campo di studi.
La microneurografia ha inoltre permesso di applicare all'uomo tecniche che finora erano riservate agli animali da esperimento e ha fatto delle percezioni del dolore un indice sicuro per valutare se una fibra sia o no capace di evocare percezioni appartenenti a questa modalità (v. Torebjörk e Hallin, 1976). Questi lavori hanno confermato la nozione fondamentale che vi sono molte fibre speciali per il dolore nel gruppo delle fibre amieliniche C e anche fra le fibre mieliniche a più lenta velocità di conduzione (gruppo III).
La microneurografia nell'uomo ha aperto un nuovo campo di ricerca nella fisiologia del dolore. La precisa indicazione percettiva renderà possibile anche la conoscenza degli stimoli che eccitano i recettori per il dolore. Tutto fa ritenere che siano di natura chimica (v. somatoestesia).
Nello studio psicofisico sulle sensazioni dolorose si è fatta distinzione fra un dolore iniziale immediato e un secondo successivo; per es., con una puntura d'ago si nota che la dolorosa trafittura iniziale è seguita da una sensazione pungente e irradiata. È stata formulata l'ipotesi che la doppia sensazione di dolore sia condotta da gruppi di fibre con due differenti velocità di conduzione. L'ipotesi è ancora controversa. Le registrazioni da singole fibre attualmente disponibili dimostrano che scariche ad alta frequenza sono seguite da scariche a frequenze più basse. Le fibre del dolore hanno campi recettivi che vanno da un paio di millimetri a ben oltre 1 centimetro di diametro lungo un asse. Se si ammette che le nostre esperienze sensitive siano dovute principalmente alle fibre dolorifiche amieliniche, sembra superfluo supporre una doppia innervazione di un punto doloroso per spiegare il primo e il secondo dolore, pur tenendo conto dell'esistenza di fibre dolorifiche fra le sottili fibre mieliniche.
Sebbene un certo numero di meccanocettori a lento adattamento rispondano al raffreddamento locale della pelle, i veri ‛recettori del freddo' sono non soltanto molto più sensibili alla stimolazione termica ma hanno inoltre un'elevata sensibilità dinamica al proprio stimolo adeguato (v. Iggo, 1969). In un primo tempo Hensel e Zotterman (v., The response of..., 1951) e Dodt e Zotterman (v., 1952) li distinsero dai meccanocettori; questi autori ottennero dalle fibre della lingua del gatto - che scaricano costantemente in risposta alle temperature riportate sull'ascissa - curve di sensibilità come quelle illustrate nella fig. 20. Molto lavoro sperimentale svolto recentemente da diversi gruppi (Burgess e Perl, Hensel, Iggo) ha esteso queste ricerche ai termocettori della pelle glabra o pelosa di diversi animali, uomo compreso.
Non sembra esservi alcuna differenza fondamentale fra i termocettori della pelle glabra o pelosa dei Primati. Iggo riporta velocità di conduzione più elevate nella pelle glabra, con medie intorno ai 9 m/s contro i 5 m/s della pelle pelosa. Le fibre di questi recettori sarebbero piccole, mieliniche del gruppo delta. Ma vi sono recettori sia del freddo sia del caldo anche nell'ambito C delle velocità di conduzione: la fig. 21 ne illustra alcuni (v. Hensel e altri, 1960). In generale, si è riscontrato che è molto più facile individuare i recettori del freddo che i recettori del caldo, il che è in accordo con le esperienze sensitive dei punti freddi e caldi riportati nella tab. II. I recettori del freddo di Iggo (v., 1969), nei Primati, avevano campi recettivi a macchie e le velocità di conduzione delle loro fibre afferenti variavano da 0,6 a 15,3 m/s, cioè da C a delta. I massimi delle curve di sensibilità dinamica e statica erano intorno a 30 °C.
I recettori del caldo, sebbene scarsi nella pelle, sembrano abbondare in regioni speciali. Gli esempi dati sono lo scroto del ratto (Iggo) e il dorso del naso del gatto (v. Hensel e Kenshalo, 1969; v. fig. 22). Un paragone fra le massime sensibilità statiche dei recettori del freddo e del caldo di questa regione dava 36 impulsi/s per i recettori del caldo a 46 °C e 9 impulsi/s per le fibre del freddo a 27 °C. I recettori del caldo avevano sensibilità dinamiche massime dell'ordine di 200 impulsi/s.
Il fatto che termocettori fondamentalmente identici si trovino nella pelle glabra o pelosa è di notevole interesse, in considerazione delle precedenti identificazioni delle sensazioni di temperatura con precise terminazioni incapsulate. Poiché queste mancano nelle zone pelose della pelle, sensibilità specifiche al caldo e al freddo non devono necessariamente essere inerenti all'incapsulamento.
d) Recettori olfattivi.
I chemocettori dell'olfatto e del gusto sono tra quelli studiati più a fondo. Come sempre, la possibilità di studio è funzione dell'accessibilità. La mucosa olfattiva dei Mammiferi non si può dire particolarmente accessibile; infatti, circa 20.000 recettori convergono verso una cellula nel bulbo olfattivo e quindi devono essere piccolissimi per trovare posto nell'epitelio nasale. Per chiarezza è dunque necessario cominciare con gli Invertebrati. La mucosa olfattiva di Molluschi mostrata nella fig. 23 contiene recettori che sono 150 volte più grandi di quelli dei Vertebrati: è stato quindi possibile introdurvi dei microelettrodi per registrare i loro potenziali di membrana. Sotto l'influenza di quantità controllate di sostanze odorose come alcool, acetato di amile, ammoniaca e benzene, immesse in correnti d'aria a velocità costanti, queste cellule rispondono con potenziali generatori (potenziali del recettore), un esempio dei quali è stato dato nella fig. 1 per illustrare la generazione di impulsi per depolarizzazione (v. Arvanitaki e altri, 1967).
Abbiamo rilevato in precedenza che un certo numero di questi recettori risponde alla stimolazione olfattiva con iperpolarizzazione, come è mostrato nella fig. 24. L'effetto dell'iperpolarizzazione è inibitorio, perciò è necessario iniziare con un recettore attivo per misurare l'effetto dell'iperpolarizzazione sull'effettivo messaggio proveniente dalla cellula, come è stato fatto nell'esperimento della fig. 24. Alcune cellule scaricano impulsi spontaneamente o possono essere depolarizzate al livello di scarica sotto l'influenza di varie miscele odorose, mentre altre sostanze possono iperpolarizzarle. Si verificheranno così combinazioni di eccitazione e inibizione che servono alla discriminazione olfattiva degli stimoli complessi. Le diverse forme dei potenziali generatori delle differenti cellule recettrici erano considerate ulteriori indicazioni per la discriminazione, ma bisogna sottolineare che parlare di ‛indicazioni' per la discriminazione significa passare dalla descrizione dei fatti alla formulazione di una ipotesi teleologica per spiegarne il significato.
D'altra parte, le funzioni dei recettori devono essere sempre interpretate nei termini dell'informazione che essi forniscono; poichè il risultato globale della stimolazione odorosa è altamente discriminativo, si è costretti a cercare indicazioni adatte alla discriminazione a tutti i livelli. In questo organismo primitivo certi recettori rispondono soltanto ad alcuni odori e non ad altri. Ancora, una data cellula reagisce sempre nella stessa maniera alla stessa molecola odorosa; ma cellule diverse possono rispondere a questa molecola con diversi tipi di potenziale del recettore. Si suppone che vi sia un numero sufficiente di possibili permutazioni entro l'insieme delle risposte di tutti i recettori olfattivi per rendere possibile e, naturalmente, anche trasmissibile, la discriminazione alla stazione successiva.
Il maschio del baco da seta (Bombyx mori) è un invertebrato particolarmente interessante per ciò che riguarda l'olfatto, perché localizza la femmina grazie a una sostanza di attrazione sessuale che questa produce in una ghiandola situata nelle pieghe intrasegmentali vicino all'estremità superiore addominale. La femmina è insensibile al proprio odore né può avvertirlo lo sperimentatore, in qualsiasi concentrazione. Gli esperimenti con questa sostanza (v. i contributi di Schneider, 1963), alla quale è stato dato il nome di bombicolo, sono stati molto facilitati dalla sua identificazione chimica, operata da Butenandt e altri (v., 1961), come esadecadien-10-trans, 12-cis-1-olo. Sostanze siffatte, che mediano l'informazione fra individui della stessa specie, sono state chiamate ‛feromoni' (v. ormoni negli invertebrati).
I recettori olfattivi degli Insetti sono situati sulle antenne entro strutture che appaiono come setole o spine di diverse forme (coni, fiaschi, cupole ecc.). In quanto tali strutture sono provviste di almeno una cellula nervosa sensitiva, esse hanno anche funzioni sensitive come chemo- e meccanocettori, alcune anche come termo- e igrocettori. Ci occuperemo in seguito di quei recettori olfattivi del baco da seta, studiati in dettaglio, che sono specificamente sensibili al bombicolo. Delle circa 40.000 fibre afferenti del nervo antennale di Rombyx, circa 15.000 trasportano informazioni su questo feromone sessuale. La fig. 25 mostra la sezione trasversale di un ramo antennale, mettendo in evidenza le apposite strutture insieme con alcuni altri organi terminali. Vi sono sensille di diversi tipi, e ciascuna sensilla consiste nel frammento morfologicamente differenziato di cuticola che contiene cellule nervose sensoriali, cellule formative e cellule accessorie. Si è trovato che le sensille basioconiche (v. fig. 25) contengono i recettori olfattivi. La loro cuticola è sottile e provvista di pori attraverso i quali l'aria è in contatto diretto con i dendriti sensoriali. È interessante notare che i maschi e le femmine dei filugelli hanno sensille identiche, e ciò nonostante solo i maschi rispondono al bombicolo. La base strutturale determinante di questo alto grado di specificità è sconosciuta. I maschi e le femmine rispondono nello stesso modo ad altri odori.
Tuttavia, la risposta del filugello al bombicolo o a frammenti della ghiandola femminile che lo contengono è la base di un metodo comportamentale di saggio, al quale è stato aggiunto un metodo diretto di registrazione del potenziale generatore depolarizzante sotto forma di reazione massiva dell'antenna (l'elettroantennogramma, EAG) o di risposta ottenuta da una singola sensilla con la tecnica microelettrodica come illustrato nella fig. 26. La soglia comportamentale è 10-11 μg di bombicolo per ml, una concentrazione incredibilmente piccola nell'aria che passa vicino alle antenne. Nel maschio di Bombyx l'elettroantennogramma in funzione della concentrazione di bombicolo segue la curva tracciata nella fig. 27. La risposta è piuttosto costante in un notevole intervallo, il che probabilmente significa che il maschio avverte soltanto la presenza della femmina senza saperla ancora localizzare. La parte di curva a rapida crescita probabilmente denota la discriminazione d'intensità necessaria a portare il maschio nella giusta direzione.
Alcune caratteristiche del recettore del bombicolo sono di interesse particolare: in primo luogo, il suo elevatissimo grado di specificità di specie; in secondo luogo, l'intervallo d'intensità a cui risponde, che è ampio quanto quello dell'occhio; ma bisogna ricordare che la retina fa uso di due recettori, bastoncelli e coni, per ottenere la straordinaria estensione della sua sfera di azione dalla semioscurità alla più brillante luce del sole. L'alta specificità di specie suggerisce possibilità pratiche di sterminio degli Insetti basate sull'impiego dei feromoni: la sostanza potrebbe essere usata per catturare i maschi, sistema già messo in pratica.
Il recettore del bombicolo sembra essere l'unico a rispondere a una sola sostanza. Negli ultimi anni il notevole lavoro sui recettori olfattivi dei filugelli e di altri insetti ha mostrato che la maggior parte delle sensille olfattive hanno spettri di risposta larghi e sovrapposti, pur avendo la massima sensibilità (o specificità) per una sostanza. Si è dimostrato inoltre che recettori singoli possono reagire con iperpolarizzazione primaria. Ciò produce inibizione della scarica in una cellula che si scarica (v. fig. 24). Non è necessario che questi recettori si scarichino spontaneamente. Le sensibilità sovrapposte agli odori garantiranno il verificarsi di combinazioni di eccitazione e inibizione, che si deve supporre possano aiutare la discriminazione olfattiva. Mentre è possibile prevedere che tali combinazioni portino a differenze nella codificazione olfattiva delle frequenze degli impulsi come complessi schemi da un gran numero di recettori, non è ancora possibile identificare alcuna risposta centrale in base al tipo della sua scarica periferica. Essere riusciti a identificare un recettore specifico, la sensilla del bombicolo, con una altrettanto specifica risposta comportamentale, è stato comunque un importante passo nella giusta direzione.
Gli esperimenti sull'olfatto degli Insetti ci hanno portato un po' più vicino alla comprensione del senso dell'olfatto nei Mammiferi, olfatto che tutti hanno avuto modo di ammirare nei cani. Il cane è stato in effetti il primo animale in cui è stata registrata una risposta elettrica massiva dell'epitelio olfattivo agli odori (v. Hosoya e Yoshida, 1937); più tardi la stessa registrazione è stata ottenuta nel coniglio (v. Ottoson, 1954). Da allora è stata osservata in un gran numero di animali, dalla rana in su nella scala zoologica. Si è pensato che si trattasse del potenziale generatore dei recettori olfattivi, anche se questi si trovano fra le cosiddette cellule di sostegno, che sono cellule gliali prive di nervi afferenti. Ultimamente si è dimostrato che anche altrove le cellule gliali sono capaci di produrre risposte elettriche, e infatti attualmente vi sono prove in favore dell'ipotesi che la componente dominante della risposta elettrica retinica alla luce (PII nella terminologia corrente) sia un potenziale gliale. Vi sono anche prove pro e contro la teoria che l'elettro-olfattogramma dei Vertebrati sia la risposta del generatore dei recettori olfattivi (v. Takagi, 1967).
Purtuttavia, l'insieme delle osservazioni indica che i meccanismi dei recettori olfattivi sono fondamentalmente simili nei Vertebrati e negli Invertebrati. Con il miglioramento delle tecniche microelettrodiche oggi è diventato possibile studiare risposte altamente localizzate anche nei Vertebrati (rane, Uccelli) ed è ormai accertato che i meccanismi recettori anche a questo livello fanno uso delle depolarizzazioni e delle iperpolarizzazioni per aumentare o diminuire la frequenza di scarica, e che, con questi mezzi, specificità sovrapposte, come si è detto sopra, producono tipi costanti, altamente differenziati, di scarica. Questi, stando alle attuali conoscenze, servono da indizi per la discriminazione olfattiva. Una rassegna su questi problemi si può trovare in T. Hayashi, 1967.
e) Controllo inibitorio centrifugo.
Abbiamo visto che i recettori sono simili ai neuroni centrali in quanto possono rispondere agli stimoli sia con una eccitazione sia con una inibizione, e abbiamo anche studiato il controllo centrifugo dei recettori da parte di meccanismi eccitatori (fusi neuromuscolari). Descriveremo in questo paragrafo un caso di controllo centrifugo dovuto a inibizione periferica, scoperto nei recettori di stiramento dei Crostacei (gamberi, aragoste). Questi organi terminali sono stati descritti da Alexandrowicz (v., 1951 e 1967). Lo studio della loro funzione, iniziato da Wiersma e altri (v., 1953), ha attirato molti ricercatori nella scia di Kuffler ed Eyzaguirre (v. Kuffler, 1958-1959), che hanno applicato per primi la tecnica microelettrodica a questi organi.
L'anatomia del recettore di stiramento dei Crostacei, situato sul lato dorsale della coda, è illustrata nella fig. 28. Vi sono speciali muscoli recettori (MR), che non si possono chiamare fibre muscolari nello stesso senso in cui il termine è usato nella fisiologia dei Vertebrati, per quanto essi consistano di miofibrille contrattili, e la striatura trasversa schematizzata nella figura può essere vista anche al microscopio. Il muscolo più profondo MR 2 ha la striatura più fitta e produce una contrazione più rapida di quello superficiale MR 1. Ciascuno dei due è provvisto di una cellula recettrice (CR) e fondamentalmente l'innervazione sia motoria che sensitiva dei due tipi di muscoli recettori ha la stessa organizzazione. Al centro del muscolo v'è una zona in cui le miofibrille sono state largamente sostituite da tessuto connettivo che forma una struttura che ricorda il tendine. In questa regione la cellula recettrice estende le sue propaggini dendritiche. Sia nell'aragosta (Homarus) sia nel gambero (Astacus) i dendriti terminano nel tessuto connettivo e non negli elementi muscolari (come si credeva una volta). I muscoli recettori hanno l'innervazione motoria in comune con i vicini muscoli estensori, quelli a lenta contrazione MR 1 con gli estensori lenti, quelli a rapida contrazione MR 2 con gli estensori rapidi. I nervi accessori sono responsabili del processo inibitorio che verrà descritto più sotto. Il nervo accessorio sottile, secondo Alexandrowicz, è ‟il più difficile da individuare fra tutti i componenti dell'ORM" (organo recettore muscolare). Nel lavoro che descriverò è probabile che fosse stato stimolato solo il nervo accessorio spesso. La fibra x della fig. 28 è un'entità sconosciuta dal punto di vista funzionale. Recenti ricerche suggeriscono che sia anch'essa inibitoria.
L'ordinamento dell'innervazione ricorda quello dei fusi neuromuscolari della rana; anche in quel caso, infatti, il muscolo intrafusale del recettore di stiramento ha alimentazione motoria comune con i muscoli extrafusali circostanti. Nei Mammiferi, però, la maggior parte delle fibre nervose che vanno alla muscolatura intrafusale è indipendente da quella dei vicini muscoli extrafusali (v. sopra, È b). In ambedue le classi di animali il recettore subisce deformazione da stiramento o contrazione, ma è probabile che nel gambero vivente la contrazione sia uno stimolo meno importante dello stiramento, anche se dovrebbe contribuire a stabilire la sensibilità dell'organo mantenendovi la tensione.
Se si inserisce un microelettrodo nel corpo del recettore e si provoca uno stiramento tirando il guscio o piegando le giunture, l'organo terminale risponde con un potenziale generatore che è rapido e fasico nel muscolo rapido (MR 2), è lento e tonico nel muscolo lento (MR 1). La fig. 29 illustra questa risposta nel muscolo lento, per un'estensione debole (A), e per un'estensione tale da generare impulsi nervosi (B). Questo potenziale del recettore deriva dai dendriti deformati dallo stiramento provocato. In questa struttura è possibile seguire il corso dei fenomeni elettrici spostando un microelettrodo lungo l'organo verso l'alto e registrare anche dal nervo afferente nel punto in cui si diparte dalla cellula sensitiva. In questo modo è stato possibile dimostrare che gli impulsi sono generati nel nervo stesso e non nell'organo terminale (v. È a).
Tuttavia il fatto nuovo e sorprendente circa il recettore di stiramento dei Crostacei è l'esistenza dei punti di contatto inibitori, o sinapsi, che sopprimono il potenziale generatore producendo ripolarizzazione o iperpolarizzazione, come è illustrato nella fig. 30. I punti di contatto sono formati dalle terminazioni dei nervi accessori. Questo è un meccanismo ben noto grazie ai numerosi studi effettuati su neuroni centrali di Vertebrati e Invertebrati, e implica un controllo, da parte di sinapsi, delle scariche neuroniche che i nervi inibitori distribuiscono al soma e ai dendriti delle cellule nervose. È ben noto che la trasmissione dell'eccitazione e dell'inibizione nelle sinapsi centrali è mediata da speciali mediatori chimici (v. sinapsi), e quindi dovrebbe essere sufficiente dire che l'inibizione centrifuga dei recettori di stiramento dei Crostacei rientra da tutti i punti di vista nel caratteristico schema illustrato dalla fisiologia generale del sistema nervoso centrale. L'accessibilità di questo recettore lo ha reso molto utile nello studio dell'inibizione sinaptica; il suo mediatore chimico sembra essere l'acido gamma-amminobutirrico, e prove sperimentali indicano che l'inibizione è dovuta a variazioni di conduttanza (aumento della permeabilità). Gli ioni implicati sono potassio e cloro. L'inibizione è del tipo conosciuto come postsinaptico.
f) Linea laterale, apparato vestibolare.
Queste terminazioni sensoriali appartengono al sistema acustico-laterale (cioè linea laterale, organi vestibolari, coclea), i cui nomi indicano una comune origine. Nel corso della differenziazione filogenetica gli organi dell'equilibrio e dell'udito si sono sviluppati da cellule ciliate situate in superficie, sensibili alle vibrazioni. È di notevole interesse generale seguire la traccia dello sviluppo della coclea (per una più ampia trattazione di questo organo, v. udito) dalla linea laterale, perché questa e gli organi vestibolari sono stati studiati a fondo dal punto di vista sia anatomico che fisiologico. La fig. 31 mostra un disegno schematico del canale della linea laterale in un pesce teleosteo (osseo). Il microscopio elettronico ha rivelato dettagli importanti della sua istologia, ma al tempo in cui Sand (v., 1937) svolgeva la sua ricerca pionieristica registrando l'attività di impulsi dai nervi della linea laterale di oltre cento esemplari di pesci d'acqua dolce e salata, si sapeva già che il canale contiene cilia sensoriali che stanno ‟come canne nel letto di un fiume". Ciò spinse Sand a cercare di perfondere il canale con un liquido. Nei pesci teleostei apparve impossibile inserire una cannula in queste delicate strutture, ma poi si riuscì a utilizzare grandi esemplari di un elasmobranchio, la razza Raja. Sand riuscì a inserire e a legare nelle due estremità dell'anello iomandibolare del canale un tubicino di vetro di 1 mm di diametro: legati i tubicini in loco, egli poté quindi perfondere il canale nelle due direzioni e registrare dal nervo corrispondente per 6-8 ore.
Nella fig. 32, tratta dal lavoro di Sand, si vede che vi è attività spontanea nei nervi della linea laterale, il che conferma precedenti osservazioni su questo organo (v. Hoagland, 1933), ma la nuova importante scoperta fu che la frequenza di scarica dai nervi stessi è aumentata o soppressa a seconda della direzione del flusso. Sand sintetizzò i suoi risultati nel modo esposto nella tab. III.

Sand riuscì anche a registrare da recettori isolati, cioè da preparati di fibre singole, e poté così verificare che effettivamente v'erano due tipi di recettori, eccitati e resi silenti dal flusso di liquido nelle opposte direzioni. ‟È evidente - affermò - che recettori antagonisti sono mescolati in un assortimento casuale in ciascun gruppo di cellule sensoriali".
Recenti ricerche (v. Flock e Wersäll, 1962; v. Flock, 1965), basate sull'impiego combinato della microscopia elettronica e dell'elettrofisiologia, hanno molto ampliato il lavoro di Sand, e anche i nuovi risultati confermano la somiglianza fondamentale della struttura e delle proprietà di tutti gli organi appartenenti al sistema acustico-laterale. Nelle forme molto primitive di pesci, gli organi terminali sono strutture epidermiche aperte e le loro cilia sensoriali sporgono direttamente nell'acqua circostante. La linea laterale completamente sviluppata (per esempio in Lota) contiene terminazioni sensoriali situate in aree distanti tra loro da 5 a 10 mm. Queste si trovano su una protuberanza a forma di sella contenente tessuto connettivo e di sostegno, compenetrato da vasi sanguigni e da fibre nervose che raggiungono le basi delle cellule ciliate. Dalla cima di ciascuna cellula ciliata il fascio protrudente delle cilia sensoriali entra in una cupola gelatinosa che quasi riempie il canale. Il piegamento della cupola è lo stimolo meccanico per il gruppo di cilia situato nel suo interno. Questa organizzazione basilare, cioè cellule sensoriali le cui cilia sono conficcate in una cupola, ricorre nell'apparato vestibolare, come si vede nella parte bassa della fig. 36. Nella coclea la cupola è sostituita da una membrana (v. udito).
Uno schema dell'organo sensoriale della linea laterale si trova nella fig. 33. Le cilia sensoriali sono di due tipi, kinocilia (nere nella figura) e stereocilia. Per ciascuna cellula ciliata v'è un kinocilium e circa 50 stereocilia, che possono essere identificate chiaramente in base alle caratteristiche strutturali svelate dal microscopio elettronico (v. Flock, 1965). Lo schema mostra che il kinocilium è alternativamente all'una e all'altra estremità del gruppo di cellule ciliate. Tutta la struttura è polarizzata funzionalmente in modo che il piegamento della cupola verso il polo kinociliare provoca una caduta di potenziale con un aumento della frequenza della scarica afferente; il piegamento nella direzione opposta, che la allontana quindi dal kinocilium, provoca un aumento del potenziale con diminuzione della frequenza della scarica afferente. Nei canali della linea laterale circa metà delle cellule ciliate hanno i poli kinociliari orientati verso la testa, e l'altra metà verso la coda (v. fig. 33). Vi sono quindi, come Sand aveva previsto, due tipi antagonisti di recettori nella stessa area recettrice.
Anni or sono Harris e collaboratori (v., 1970) sono riusciti a registrare intracellularmente da cellule ciliate ben individuate di Necturus maculosus durante la stimolazione di singole cupole a 27 Hz con una pipetta vibrante di 10 μm di diametro. In questi esperimenti è stato possibile osservare che, in cellule ciliate differenti, i potenziali del recettore erano sfasati di 180°, come previsto. Per il momento non si sa come questi potenziali del recettore attivino le sinapsi per mezzo delle quali i nervi afferenti sono in contatto con le cellule ciliate (v. fig. 33), né si conosce il meccanismo che genera attività spontanea. Sand osservò che questa è puramente neurogena e non causata da movimento del liquido, vibrazione o pressione dall'esterno.
Nel 1930 Wever e Bray scoprirono il cosiddetto potenziale microfonico della coclea, che riproduce con alto grado di fedeltà onde sinusoidali e altre funzioni d'onda del campo acustico. In base al presupposto che la coclea è una differenziazione filogenetica della linea laterale, ci si aspetterebbe di trovare potenziali elettrici microfonici anche in quest'ultima. Poiché il potenziale microfonico è una risposta massiva generata dalle cellule ciliate, e dato che quelle della linea laterale - come abbiamo spiegato prima - hanno una doppia polarità, ci si potrebbe aspettare che le due direzioni delle kinocilia siano riprodotte come un raddoppiamento della frequenza della risposta microfonica a un'onda sinusoidale. È noto da molto tempo che il potenziale microfonico della linea laterale ha una frequenza doppia di quella di uno stimolo vibratorio.
Spesso la conoscenza dello stimolo adeguato di un organo di senso permette allo sperimentatore di dedurre che tipo di compiti questo svolge nell'organismo cui appartiene. Nel caso della linea laterale s'incontrano difficoltà che non si possono dire completamente superate. Questo meccanocettore è sensibile al tatto, alla vibrazione, alla pressione, in breve a qualsiasi cosa che faccia premere l'endolinfa del canale sulla cupola. Non si può evitare la stimolazione della cupola se la pelle del pesce per caso entra in contatto con un oggetto solido; Sand ha visto il nervo laterale scaricare per la contrazione sporadica di poche fibre muscolari di una pinna e scaricare in modo imponente quando il corpo del pesce si flette nuotando. Ciò nonostante, non si tratta probabilmente di un sostituto propriocettivo dei recettori dei muscoli, perché i muscoli sono provvisti di propri recettori e lo stesso ragionamento, mutatis mutandis, è valido anche per la pelle. In questi casi si può solo supporre che la linea laterale fornisca informazioni supplementari.
Importanti argomenti sono stati addotti da van Bergeijk (v., 1967) per l'interpretazione di almeno una funzione fondamentale della linea laterale. Il suo ragionamento si basa sulla sensibilità alla vibrazione che questa ha in comune con la coclea. Invece di uno stimolo acustico in campo vicino, nell'aria, quale può essere la rottura di un ramoscello, si dovrebbe considerare una corrispondente perturbazione idrodinamica nell'acqua. Questa può essere provocata dal passaggio di un altro organismo, che nuoti o che comunque si muova. La fig. 34 mostra come un rivelatore di spostamento, qual è la linea laterale, funzioni a una certa distanza, breve in paragone all'udito, ma senza richiedere un contatto effettivo. L'estensione della linea laterale lungo il corpo insieme con il carattere direzionale dei suoi segnali dovrebbe fornire una componente pure direzionale al rivelamento delle onde idrodinamiche in campo vicino. In effetti, Pesci e Anfibi sono capaci di localizzare oggetti in movimento a qualche distanza.
Le osservazioni di Sand indicano che il canale della linea laterale sarebbe un utilissimo autocontrollo per il pesce che si muove. A ogni flessione del suo corpo si genereranno asimmetrie bilaterali nel flusso dell'endolinfa attraverso i canali aperti, provocate anche dai suoi atti muscolari, per esempio quando il pesce gira da destra a sinistra. Muovendosi nell'acqua, che è un mezzo omogeneo. il pesce dovrebbe aver bisogno di informazioni direzionali di questo tipo. I recettori della linea laterale sulla testa dell'animale dovrebbero essere molto meno sensibili a stimoli come il girare a destra o a sinistra, ma la loro scarica può certo servire come un punto di riferimento per i recettori dei canali del corpo.
È interessante notare che si ritiene che gli organi vestibolari si siano sviluppati nella linea laterale della testa per chiusura dei tubi e migrazione dei nuovi canali modificati nelle strutture ossee. Questo fatto li ha liberati dall'influenza perturbante degli stimoli per i quali una ubicazione superficiale nella pelle era biologicamente utile, mentre nello stesso tempo la chiusura dei canali ha reso l'organo sensibile all'accelerazione. L'ulteriore sviluppo dei canali semicircolari in tre piani (v. fig. 35) li ha resi sensibili ad accelerazioni angolari in tutte le direzioni. Questo sviluppo ebbe luogo ai primordi dell'evoluzione. I Teleostei hanno avuto un organo vestibolare completamente sviluppato per milioni di anni, come dimostrano anche i più antichi reperti paleontologici, e non è mai stato possibile osservare l'inizio dello sviluppo sopra descritto. I ragionamenti sull'evoluzione sono congetture, ma il fisiologo rimane con un risultato chiaro e molto suggestivo: vi sono canali, cupole, cellule ciliate, kinocilia e stereocilia e fondamentali somiglianze delle scariche degli impulsi in tutti gli organi vestibolari.
Gli organi vestibolari e la coclea sono rinchiusi nel labirinto osseo, da cui li separano la perilinfa e il labirinto membranoso (v. fig. 35). Si tratta di un complesso sistema di canali e sacchi contenenti l'endolinfa. Nei Pesci vi sono tre sacchi, utriculus, sacculus e lagena, e quest'ultima è destinata a svilupparsi nella coclea della fig. 35. Nei Rettili superiori e negli Uccelli la lagena è già leggermente ritorta ed è presente un apparato uditivo. Il prodotto finale dell'evoluzione è la coclea dei Mammiferi con il suo organo del Corti (v. udito). Nei Mammiferi, quindi, solo due sacchi, utriculus e sacculus, rimangono a svolgere le loro funzioni originali.
Il dotto endolinfatico, che nei Pesci inferiori è ancora aperto e in alcuni Elasmobranchi serve a trasportare particelle di sabbia nel sacculus, si chiude e forma un sacco endolinfatico a uno stadio evolutivo ancora molto basso. I canali semicircolari esterni generalmente mancano nei Ciclostomi, ma due di essi appaiono in Petromyzon (lampreda). I Teleostei hanno un apparato vestibolare ben sviluppato con tre canali semicircolari; i loro sacculus e utriculus per lo più contengono grandi concrezioni, chiamate ‛otoliti'. In un elasmobranchio si trova solo sabbia. Nella maggior parte dei Vertebrati vi sono ‛otoconi', piccoli cristalli di calcite e aragonite, adesi a una membrana che in questi organi ha sostituito la cupola della linea laterale originale. Le membrane cosi appesantite di queste strutture sono in contatto con le cilia della macula, e costituiscono un apparato con una massa sufficiente per renderlo sensibile alla gravità.
I tre canali semicircolari - il laterale (orizzontale), il posteriore e il superiore (v. fig. 35) - sono disposti su tre piani ad angolo retto fra loro. Tutti i canali terminano nell'utriculus e a una delle estremità si allargano in ampolle. Le cellule ciliate sensoriali sono disposte su una cresta interna chiamata crista ampullaris. Le cilia si allungano fin dentro la cupola, come è mostrato nella fig. 36. Come nella linea laterale, lo stimolo adeguato per le cellule ciliate è lo spostamento della cupola.
Le cellule ciliate della cresta ampollare hanno subito un'ulteriore differenziazione nei Mammiferi. Con la microscopia elettronica se ne possono distinguere due tipi, ma data la mancanza di precise informazioni fisiologiche circa il significato di questa differenziazione si rimanda ai lavori originali sull'argomento (v. Wersäll e Flock, 1964). Le cilia di ciascuna cellula ciliata sono organizzate come nella linea laterale, un kinocilium e diverse stereocilia, come si vede nella fig. 37 che, confrontata con la fig. 33, rivela però un'importante differenza di orientamento: nella cresta ampollare tutte le kinocilia si trovano alla stessa estremità della cellula in modo che la risposta di tutte le cellule ciliate alla flessione della cupola è uniforme. Così, per esempio, uno spostamento utriculopeto della cupola nel canale orizzontale porta a una depolarizzazione con un aumento di scarica nel nervo; uno spostamento utriculofugo a un'iperpolarizzazione e a un'inibizione della scarica. Negli altri canali semicircolari le kinocilia sono situate verso il lato del canale delle loro creste. Poiché la posizione del kinocilium determina la direzione dell'eccitazione (v. Loewenstein e Wersäll, 1959), questi canali saranno eccitati da uno spostamento della cupola nella direzione utriculofuga. È stato recentemente dimostrato che le terminazioni ampollari sono sotto controllo efferente, probabilmente dello stesso tipo di quello dell'organo del Corti e della retina, ma il significato di questo meccanismo nella fisiologia vestibolare è ancora da chiarire.
Lo stimolo adeguato per i tre canali semicircolari è la rotazione della testa nei loro rispettivi piani. L'inerzia fa restare indietro l'endolinfa nella fase iniziale della rotazione. Ciò produce un movimento dell'endolinfa che trascina con sé la cupola nella direzione opposta alla rotazione. Il tipo di movimento a porta oscillante della cupola è stato effettivamente osservato nei Pesci (v. Steinhausen, 1933; v. Dohlman, 1941). Durante una rotazione continuata l'elasticità della cupola la fa ritornare nella posizione di riposo (in circa 30 secondi). Tali proprietà rendono questo organo di senso sensibile all'accelerazione piuttosto che alla velocità. Alla cessazione della rotazione il flusso inverte la sua direzione. I segnali positivi e negativi della scarica sono elaborati sullo sfondo di un'attività spontanea di impulsi, secondo uno schema già visto per la linea laterale (v. fig. 32). Dato che l'apparato vestibolare è simmetricamente bilaterale, l'informazione fornita al sistema nervoso centrale sarà specificata da un differenziale biauricolare. Considerando per esempio la cresta ampollare orizzontale, l'endolinfa fluirà in direzioni opposte nel canale sinistro e destro quando la testa ruota nel suo piano. Il segnale sarà positivo (+) da un lato e negativo (−) dall'altro.
Loewenstein e Sand (v., 1940) hanno riassunto i loro esperimenti di registrazione dal nervo vestibolare (mostrato nella fig. 35) nel modo illustrato nella tab. IV, che dimostra quanto siano dettagliate le indicazioni di questo analizzatore dello spazio, quando affronta il compito di misurare l'accelerazione angolare durante i movimenti della testa.

Tuttavia, lo spazio richiede anche un controllo statico delle inclinazioni costanti della testa. Serviranno a questo scopo i muscoli del collo che sono estremamente ricchi di fusi neuromuscolari e possiedono anche organi tendinei. Ma nelle macule dell'utriculus e del sacculus vi sono membrane fornite di concrezioni, abbastanza pesanti da rispondere agli stimoli gravitazionali o alle accelerazioni lineari. Nei Pesci è facile dimostrare che le membrane sono sensibili alle vibrazioni. Esse sono grandi e accessibili, ma poiché si trovano dentro la testa e sono in risonanza con alta selettività per i loro otoliti oscillanti, difficilmente la loro sensibilità alla vibrazione può essere funzionalmente importante. Per la loro eccitazione occorrerebbero grandi ampiezze di stimolazione, molto al di sopra delle soglie delle aree sensoriali della linea laterale. D'altronde nei Mammiferi v'è l'organo del Corti specializzato nella registrazione delle vibrazioni provenienti da lontano. Perciò, tutto sommato, il sacculus e l'utriculus possono essere trascurati come organi sensibili alle vibrazioni. Queste due strutture sono effettivamente organizzate al fine della sensibilità morfologica alla direzione, che abbiamo già preso in considerazione descrivendo la linea laterale e i canali semicircolari. Le kinocilia nel sacculus e nell'utriculus sono distribuite in direzioni differenti nelle differenti porzioni delle loro macule, le quali si piegano a loro volta in direzioni diverse. Per schematizzare si può dire che il sacculo registra le inclinazioni laterali della testa, l'utricolo le inclinazioni in avanti e indietro. Come altri organi di senso che misurano variazioni statiche, questi recettori presentano una scarica spontanea. Come nella linea laterale e nei canali, essi segnalano la direzione con più e meno. La fig. 38, presa da un lavoro di Adrian (v., 1943), mostra la frequenza degli impulsi in funzione dell'inclinazione della testa del gatto.
Per circa un secolo l'apparato vestibolare è stato conosciuto come l'organo dell'equilibrio (v. Camis, 1930). Questa interpretazione fu la conseguenza dei primi esperimenti di estirpazione di questi organi, che causava cambiamenti drastici nella postura e nell'andatura. Quando le tecniche di stimolazione furono perfezionate da Ewald (v., 1887), il quale applicava una pressione direttamente sul canale membranoso attraverso perforazione della parete ossea, cominciò un'era di stimolazione rigorosa. Da allora ebbe inizio una seria analisi sperimentale degli stimoli adeguati e dei loro numerosi effetti riflessi, effettuata sia nei laboratori di fisiologia, sia nelle cliniche. Ai clinici si devono gli importanti studi sul nistagmo, sui movimenti oculari riflessi alla rotazione. Fondamentalmente le risposte riflesse vestibolari dei muscoli oculari servono a mantenere costante l'orientamento dello sguardo, compensando i movimenti della testa. Similmente i numerosi riflessi degli arti e del tronco servono a mantenere l'equilibrio del corpo. Contributi importanti nell'era preelettronica allo studio dei riflessi posturali furono quelli di Sherrington (v., 1906), al volgere del secolo, e quelli di Magnus e de Kleijn (1919-1920; v. Magnus, 1924). Si stabilì la funzione del cervelletto con i suoi nuclei e quella dei muscoli del collo in collaborazione con l'apparato vestibolare. Un'idea del controllo muscolare esercitato dagli organi vestibolari si può desumere dai comuni studi clinici del nistagmo termico effettuati da Barany, intorno al 1910 (v. Camis, 1930). Le correnti convettive causate, per esempio, dall'acqua fredda iniettata in un orecchio influenzano il flusso dell'endolinfa e di conseguenza provocano nistagmo, vertigini e tendenza a cadere. Nell'era presente il volo e i viaggi spaziali hanno conferito nuova attualità a questo campo.
Bibliografia.
Adrian, E. D., The basis of sensation, London 1928.
Adrian, E. D., Mechanisms of nervous action, London 1932.
Adrian, E. D., Discharges from vestibular receptors in the cat, in ‟Journal of physiology", 1943, CI, pp. 389-407.
Adrian, E. D., Cattell, McK., Hoagland, H., Sensory discharges in single cutaneous nerve fibres, in ‟Journal of physiology", 1931, LXXII, pp. 377-404.
Alexandrowicz, J. S., Muscle receptor organs in the abdomen of Homarus vulgaris and Palinurus vulgaris, in ‟Quarterly journal of microscopical science", 1951, XCII, pp. 163-199.
Alexandrowicz, J. S., Receptor organs in thoracic and abdominal muscles of Crustacea, in ‟Biological reviews", 1967, XLII, pp. 288-326.
Arvanitaki, A., Takeuchi, H., Chalazonitis, N., Specific unitary osmoreceptor potentials and spiking patterns from giant nerve cells, in Olfaction and taste (a cura di T. Hayashi), vol. II, Oxford 1967, pp. 573-598.
Bergeijk, W. A. van, The evolution of Vertebrate hearing, in Contributions to sensory physiology (a cura di W. D. Neff), vol. II, New York 1967, pp. 1-49.
Bessou, P., Perl, E. R., Response of cutaneous sensory units with unmyelinated fibers to noxious stimuli, in ‟Journal of neurophysiology", 1969, XXXII, pp. 1025-1043.
Blix, M., Experimentel a bidrag till lösning af frågan om hudnervernas specifika energi, in ‟Uppsala läkareförenings förhandlingar", 1882-1883, XVIII, pp. 87-102, 427-440.
Boyd, I. A., Davey, M. R., Composition of peripheral nerves, Edinburgh 1968.
Brown, A. G., Iggo, A., Cutaneous receptors and afferent fibres in cat and rabbit, in ‟Journal of physiology", 1967, CXCIII, pp. 707-733.
Bullock, T. H., Horridge, G. A., Structure and function in the nervous systems of Invertebrates, vol. I, San Francisco 1965.
Burgess, P. R., Petit, D., Warren, R. M., Receptor types in cat hairy skin supplied by myelinated fibers, in ‟Journal of neurophysiology", 1968, XXXI, pp. 833-848.
Butenandt, A., Hecker, E., Hopp, M., Koch, W., Konstitutionsermittlung und Synthese des Bombykols des Sexuallockstoffes des Seidenspinners Bombyx mori, in ‟Sizungsberichte der Bayerischen Akademie der Wissenschaften zu München. Mathematisch-physikalische Klasse", 1961.
Camis, M., The physiology of the vestibular apparatus (a cura di R. S. Creed), Oxford 1930.
Crowe, A., Matthews, P. B. C., The effects of stimulation of static and dynamic fusimotor fibres on the response to stretching of the primary endings of muscle spindles, in ‟Journal of physiology", 1964, CLXXIV, pp. 109-131.
Dodt, E., Zotterman, Y., Mode of action of warm receptors, in ‟Acta physiologica scandinavica", 1952, XXVI, pp. 346-357.
Dohlman, G., Die Rolle der Perilymphe bei den vestibulären Reaktionen, in ‟Archiv für Ohren-, Nasen- und Kehlkopfheilkunde", 1941, CL, pp. 25-30.
Douglas, W. W., Ritchie, J. M., Non-medullated fibres in the saphenous nerve which signal touch, in ‟Journal of physiology", 1957, CXXXIX, pp. 385-399.
Douglas, W. W., Ritchie, J. M., Straub, R. W., The role of nonmyelinated fibres in signalling cooling of the skin, in ‟Journal of physiology", 1960, CL, pp. 266-283.
Eccles, J. C., The physiology of nerve cells, Baltimore 1957.
Eldred, E., Granit, R., Merton, P. A., Supraspinal control of the muscle spindles and its significance, in ‟Journal of physiology", 1953, CXXII, pp. 498-523.
Erlanger, J., Gasser, H. S., Electrical signs of nervous activity, Philadelphia 1937.
Euler, C. von, Selective responses to thermal stimulation of mammalian nerves, in ‟Acta physiologica scandinavica", 1947, XIV, suppl. 45.
Ewald, J. R., Zur Physiologie der Bogengänge, in ‟Pflügers Archiv", 1887, XLI, pp. 463-483.
Eyzaguirre, C., Kuffler, S. W., Processes of excitation in the dendrites and in the soma of single isolated sensory nerve cells of the lobster and crayfish, in ‟Journal of general physiology", 1955, XXXIX, pp. 87-119.
Fechner, G. T., Elemente der Psychophysik, Leipzig 1962.
Flock, Å., Electron microscopic and electrophysiological studies on the lateral line organ, in ‟Acta oto-laryngologica", 1965, suppl. 199.
Flock, Å., Wersäll, J., A study of the orientation of the sensory hairs of the receptor cells in the lateral line organ of fish, with special references to the function of the receptors, in ‟Journal of cell biology", 1962, XV, pp. 19-27.
Flourens, M., Expériences sur les canaux semi-circulaires de l'oreille dans les Mammifères, in ‟Mémoires de l'Académie Royale des Sciences, Paris", 1830, IX, pp. 466-477.
Frisch, K. von, Die Sonne als Kompass im Leben der Bienen, in ‟Experientia", 1950, VI, pp. 210-221.
Gammon, B. D., Bronk, D. W., The discharge of impulses from Pacinian corpuscules in the mesentery and its relation to vascular changes, in ‟American journal of physiology", 1935, CXIV, pp. 77-84.
Granit, R., Sensory mechanisms of the retina, London 1947; reprint New York 1963.
Granit, R., Receptors and sensory perception, New Haven 1955.
Granit, R. (a cura di), Muscular afferents and motor control. Nobel Symposium I., Stockholm 1966.
Granit, R., The basis of motor control, London 1970.
Granit, R., Holmgren, B., Merton, P. A., The two routes for excitation of muscle and their subservience to the cerebellum, in ‟Journal of physiology", 1955, CXXX, pp. 213-224.
Granit, R., Kernell, D., Lamarre, Y., Algebrical summation in synaptic activation of motoneurones firing within the ‛primary range' to injected currents, in ‟Journal of physiology", 1966, CLXXXVII, pp. 379-399.
Hagbarth, K.-E., Microneurography in man, in Sensory functions of the skin in Primates (a cura di Y. Zotterman), Oxford 1976, pp. 129-136.
Hagbarth, K.-E., Vallbo, Å. B., Single unit recordings from muscle nerves in human subjects, in ‟Acta physiologica scandinavica", 1969, LXXVI, pp. 321-334.
Hallin, R. G., Torebjörk, H. E., Studies on cutaneous A and C fibre afferents, skin nerve blocks and perception, in Sensory functions of the skin in Primates (a cura di Y. Zotterman), Oxford 1976, pp. 137-149.
Hardy, M., Observations on the innervation of the macula sacculi in man, in ‟Anatomical record", 1934, LIX, pp. 403-418.
Harris, G. G., Frishkopf, L. S., Flock, A., Receptor potentials from hair cells of the lateral line, in ‟Science", 1970, CLXVII, pp. 76-79.
Hayashi, T. (a cura di), Olfaction and taste, vol. II, Oxford 1967.
Hensel, H., Iggo, A., Witt, I., A quantitative study of sensitive cutaneous thermoreceptors with C afferent fibres, in ‟Journal of physiology", 1960, CLIII, pp. 113-126.
Hensel, H., Kenshalo, D. R., Warm receptors in the nasal region of cats, in ‟Journal of physiology", 1969, CCIV, pp. 99-112.
Hensel, H., Zotterman, Y., Quantitative Beziehungen zwischen der Entladung einzelner Kältefasern und der Temperatur, in ‟Acta physiologica scandinavica", 1951, XXIII, pp. 291-319.
Hensel, H., Zotterman, Y., The response of mechanoreceptors to thermal stimulation, in ‟Journal of physiology", 1951, CXV, pp. 16-24.
Hoagland, H., Electrical responses from the lateral line nerves of catfish. I., in ‟Journal of general physiology", 1933, XVI, pp. 695-714.
Hosoya, Y., Yoshida, H., Über die bioelektrische Erscheinungen an der Riechschleimhaut, in ‟Japanese journal of medical science. Biophysics", 1937, V, pp. 22-23.
Iggo, A., Single C fibres from cutaneous receptors, in ‟Journal of physiology", 1958, CXLIII, pp. 47P-48P.
Iggo, A., Cutaneous thermoreceptors in Primates and sub-primates, in ‟Journal of physiology", 1969, CC, pp. 403-430.
Iggo, A., Muir, A. R., The structure and function of a slowly adapting touch corpuscle in hairy skin, in ‟Journal of physiology", 1969, CC, pp. 763-796.
Jansen, J. K. S., Wallöe, L., Transmission of signals from muscle stretch receptors to the dorsal spinocerebellar tract, in The cerebellum in health and disease (a cura di W. S. Field e W. D. Willis), St. Louis 1970, pp. 143-171.
Katz, B., Depolarization of sensory terminals and the initiation of impulses in the muscle spindle, in ‟Journal of physiology", 1950, CXI, pp. 261-282.
Kernell, D., Input resistance, electrical excitability, and size of ventral horn cells in the cat spinal cord, in ‟Science", 1966, CLII, pp. 1637-1640.
Knibestöl, M., Vallbo, Å. B., Single unit analysis of mechanoreceptor activity from the human glabrous skin, in ‟Acta physiologica scandinavica", 1970, LXXX, pp. 178-195.
Kolmer, W., Der Bau der Endapparate des Nervus octavus und deren physiologische Bedeutung, in ‟Ergebnisse der Physiologie", 1911, XI, pp. 372-416.
Kries, J. von, Allgemeine Sinnesphysiologie, Leipzig 1923.
Kuffler, S. W., Excitation and inhibition in single nerve cells, in ‟Harvey lectures", 1958-1959, LIV, pp. 176-218.
Kuffler, S. W., Eyzaguirre, C., Synaptic inhibition in an isolated nerve cell, in ‟Journal of general physiology", 1955, XXXIX, pp. 155-184.
Leksell, L., The action potential and excitatory effects of the small ventral root fibres to skeletal muscle, in ‟Acta physiologica scandinavica", 1945, X, suppl. 31.
Liddell, E. G. T., Sherrington, C. S., Reflexes in response to stretch (myotatic reflexes), in ‟Proceedings of the Royal Society" (Series B), 1924, XCVI, pp. 212-242.
Lloyd, D. P. C., Neuron patterns controlling transmission of ipsilateral hind limb reflexes in cat, in ‟Journal of neurophysiology", 1943, VI, pp. 293-314.
Loewenstein, O., Sand, A., The individual and integrated activity of the semicircular canals of the Elasmobranch labyrinth, in ‟Journal of physiology", 1940, XCIX, pp. 89-101.
Loewenstein, O., Wersäll, J., A functional interpretation of the electronmicroscopic structure of the sensory hairs in the cristae of the Elasmobranch Raja clavata in terms of the directional sensitivity, in ‟Nature", 1959, CLXXXIV, pp. 1807-1808.
Loewenstein, W. R., Generator processes of repetitive activity in a Pacinian corpuscle, in ‟Journal of general physiology", 1958, XLI, pp. 825-845.
Loewenstein, W. R., Mendelson, M., Components of receptor adaptation in a Pacinian corpuscle, in ‟Journal of physiology", 1965, CLXXVII, pp. 377-397.
Loewenstein, W. R., Skalak, R., Mechanical transmission in a Pacinian corpuscle. An analysis and a theory, in ‟Journal of physiology", 1966, CLXXXII, pp. 346-378.
Magnus, R., Körperstellung, Berlin 1924.
Matthews, P. B. C., The differentiation of two types of fusimotor fibre by their effects on the dynamic response of muscle spindle primary endings, in ‟Quarterly journal of experimental physiology", 1962, XLVII, pp. 324-333.
Matthews, P. B. C., Muscle spindles and their motor control, in ‟Physiological reviews", 1964, XLIV, pp. 219-288.
Mountcastle, V. B., The neural replication of sensory events in the somatic afferent system, in ‟Pontificiae Academiae Scripta", 1965, XXX, pp. 127-169.
Müller, J., Handbuch der Physiologie des Menschen, vol. II, Coblenz 1838.
Ottoson, D., Sustained potentials evoked by olfactory stimulation, in ‟Acta physiologica scandinavica", 1954, XXXII, pp. 384-386.
Retzius, G., Biologische Untersuchungen, 4 voll., Stockholm 1892.
Reuck, A. V. S. de, Knight, J. (a cura di), Touch, heat and pain. Ciba Foundation Symposia, London 1966.
Sand, A., The mechanism of the lateral sense organs of Fishes, in ‟Proceedings of the Royal Society" (Series B), 1937, CXXIII, pp. 472-495.
Schneider, D., Electrophysiological investigation of Insect olfaction, in 1st International Symposium on olfaction and taste (a cura di Y. Zotterman), Oxford 1963, pp. 85-103.
Schneider, D., Vergleichende Rezeptorphysiologie am Beispiel der Riechorgane von Insekten, in ‟Jahrbuch der Max-Planck-Gesellschaft zur Förderung der Wissenchaften", 1963, pp. 150-179.
Schneider, D., Kaissling, K.-E., Der Bau der Antenne des Seidenspinners Bombyx mori, L. III. Das Bindegewebe und das Blutgefäss, in ‟Zoologische Jahrbücher. Anatomie", 1959, LXXVII, pp. 111-132.
Sherrington, C. S., The integrative action of the nervous system, New Haven 1906.
Steinhausen, W., Über die Beobachtung der Cupula in den Bogengangsampullen des Labyrinths des lebenden Hechts, in ‟Pflügers Archiv", 1933, CCXXXII, pp. 500-512.
Takagi, S. F., Are EOG's generator potentials?, in Olfaction and taste (a cura di T. Hayashi), vol. II, Oxford 1967, pp. 167-179.
Torebjörk, H. E., Hallin, R. G., Skin receptors supplied by unmyelinated (C) fibres in man, in Sensory functions of the skin in Primates (a cura di Y. Zotterman), Oxford 1976, pp. 457-487.
Vallbo, Å. B., Slowly adapting muscle receptors in man, in ‟Acta physiologica scandinavica", 1970, LXXVIII, pp. 315-333.
Weddell, G., Pallie, W., Palmer, E., The morphology of peripheral nerve terminations in the skin, in ‟Quarterly journal of microscopical science", 1954, XCV, pp. 483-501.
Wersäll, J., Flock, Å., Physiological aspects on the structure of vestibular end organs, in ‟Acta oto-laryngologica", 1964, LVIII, suppl. 192, pp. 85-89.
Wever, E. G., Bray, C. W., Action currents in the auditory nerve in response to acoustical stimulation, in ‟Proceedings of the National Academy of Sciences", 1930, XVI, pp. 344-350.
Wiersma, C. A. G., Furshpan, E., Florey, E., Physiological and pharmacological observations on muscle receptor organs of the crayfish, Cambarus Clarkii Girard, in ‟Journal of experimental biology", 1953, XXX, pp. 136-150.
Zotterman, Y., Touch, pain and tickling: an electrophysiological investigation on cutaneous sensory nerves, in ‟Journal of physiology", 1939, XCV, pp. 1-28.
Zotterman, Y., Thermal sensations, in Handbook of physiology, section I, Neurophysiology (a cura di H. W. Magoun), vol. I, Washington 1959, pp. 431-458.
Zotterman, Y. (a cura di), Sensory functions of the skin in Primates, Oxford 1976.
© Istituto della Enciclopedia Italiana - Riproduzione riservata


