Struttura e funzione della corteccia: circuiti, aree e comportamento
Frontiere della Vita (1999)
Struttura e funzione della corteccia: circuiti, aree e comportamento
Le interazioni corticali sono un aspetto fondamentale dell'organizzazione del cervello e sono state associate tradizionalmente ai processi cognitivi superiori. Nella prima parte di questo saggio riassumeremo brevemente le componenti di base della struttura della corteccia, passeremo in rassegna alcuni fra i diversi tipi di connessioni che mediano le interazioni corticali e presenteremo esempi dei metodi sperimentali utilizzati nello studio delle loro funzioni. Nella seconda parte discuteremo diverse proposte teoriche che mettono in relazione l'architettura neuronale e quella cognitivo a livello di sistemi su larga scala. La relazione tra struttura neuronale e comportamento non è necessariamente diretta e lo sua comprensione rimane una sfida importante per lo ricerca futura.
Introduzione
La struttura e le funzioni della corteccia sono molto complesse e dunque non è sorprendente che, per molti aspetti, stiamo appena cominciando a capire le relazioni che intercorrono tra loro. Da un punto di vista storico, il dibattito ha visto contrapporsi due opposte tendenze: il punto di vista dei localizzazionisti, che suggerisce una relazione diretta e puntuale tra funzioni o attributi specifici e aree corticali, e il punto di vista 'equipotenziale' o della azione di massa secondo il quale, invece, una data funzione non è legata a un'area specializzata, ma piuttosto è il risultato dell'attività congiunta di molteplici aree. Entrambi questi punti di vista sembrano ormai, nella loro inflessibile formulazione, una grossolana semplificazione.
Le recenti tecniche di ricerca, come gli studi di visualizzazione funzionale condotti su esseri umani mediante tomografia a emissione di positroni (PET, Positron Emission Tomography) o risonanza magnetica funzionale (fMRl, functional Magnetic Resonance Imaging), mostrano nuovi e affascinanti orizzonti relativi ai pattern di attività corticale. Questi studi mostrano in modo coerente che anche stimoli relativamente semplici portano a focolai multipli di attività distribuiti nel cervello. Dai tradizionali tentativi di localizzare l'attività psicologica, l'attenzione si è spostata al problema di identificare i sistemi neuronali e le loro reti di connessioni e a quello di capire come questi sistemi possano essere legati a certi tipi di operazioni cognitive. In alcuni casi il collegamento tra la struttura neuronale e il comportamento può essere sorprendentemente diretto. Per esempio, la mielinizzazione delle connessioni principali delle aree motorie è strettamente legata alla fase dello sviluppo in cui i bambini cominciano a camminare. Gli animali che hanno la capacità di articolare indipendentemente le dita (alto grado di 'destrezza') presentano abbondanti connessioni dirette tra i neuroni della corteccia motoria e i motoneuroni del midollo spinale. Diversi comportamenti specifici di una specie (come la cattura di una preda o la sua fuga) sono stati analizzati nei vertebrati inferiori al livello di circuiti e reti. Nel caso di funzioni più astratte come percezione, memoria o linguaggio è più difficile individuare la corrispondenza tra struttura neuronale e comportamento. Come sono collegati i diversi domini e livelli di conoscenza alle operazioni neuronali nello spazio e nel tempo? Tradizionalmente, studi complementari condotti su animali ed esseri umani hanno consentito lo studio delle componenti costitutive dell'organizzazione cerebrale e delle loro interazioni. l lavori sugli animali, in cui è più facile manipolare le condizioni sperimentali, permettono una mappatura ad alta risoluzione delle connessioni tra insiemi di neuroni e una loro caratterizzazione funzionale. Spesso i risultati possono essere ricondotti a quelli degli studi comportamentali, sebbene negli animali sia difficile (per esempio, nel caso della memoria) o addirittura impossibile (per il calcolo o per il linguaggio) prendere in considerazione le funzioni corti cali superiori. Negli esseri umani la neuropsicologia sperimentale e quella clinica hanno sollevato questioni sofisticate a livello comportamentale e cognitivo, ma in questo caso i dati anatomici e fisiologici sono necessariamente più limitati. L'importanza di combinare i punti di forza della ricerca animale con quelli della ricerca sugli esseri umani è ovvia, e tale obiettivo è stato in parte raggiunto per mezzo di un dialogo costruttivo e di strategie ben coordinate. Grazie ai continui progressi tecnologici, sicuramente vi sarà un ulteriore avvicinamento e una mutua stimolazione tra i due campi di ricerca.
Questo saggio presenterà, nella sua parte iniziale, una panoramica sugli elementi di base e sulle connessioni dell'organizzazione corticale. In questa parte, ci riferiremo principalmente ai lavori condotti su animali e forniremo esempi funzionali al livello del singolo neurone. Lo scopo non è tanto quello di proporre dei collegamenti concreti tra funzioni corti cali superiori e operazioni neuronali, quanto piuttosto quello di dare un'idea della ricchezza del repertorio anatomico che supporta tali operazioni e della complessità delle interazioni che potrebbero risultarne. Le interazioni corticali sono di importanza centrale per le funzioni e l'organizzazione del cervello, e fornirne una spiegazione risulta pertanto di primaria importanza. Quali aree sono interconnesse tra loro? Quali sono i parametri numerici, cioè quante e quali cellule in una certa area sono connesse a quante e quali in un'altra area? Quale funzione svolgono le connessioni? La ricerca in corso è ancora per lo più in una fase analitica, ma sono stati proposti diversi concetti che riassumono l'organizzazione su larga scala della corteccia, sulla base di questi e altri dati. Nella seconda parte del saggio, forniremo una breve descrizione delle idee attuali e dei tentativi di collegare questi concetti riassuntivi ad alcuni aspetti dei dati sulla connettività.
Le aree corticali
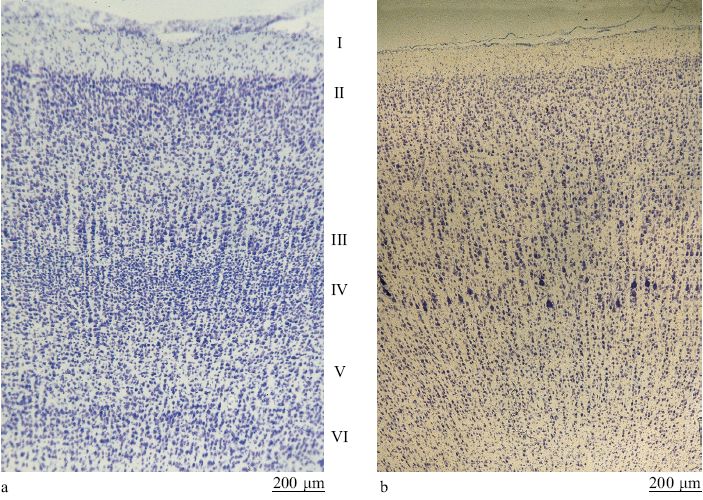
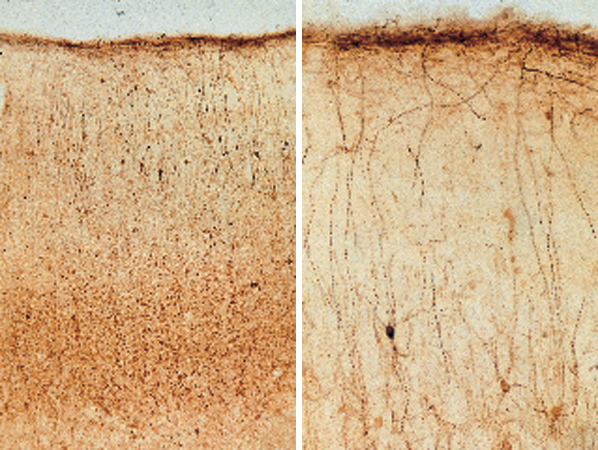
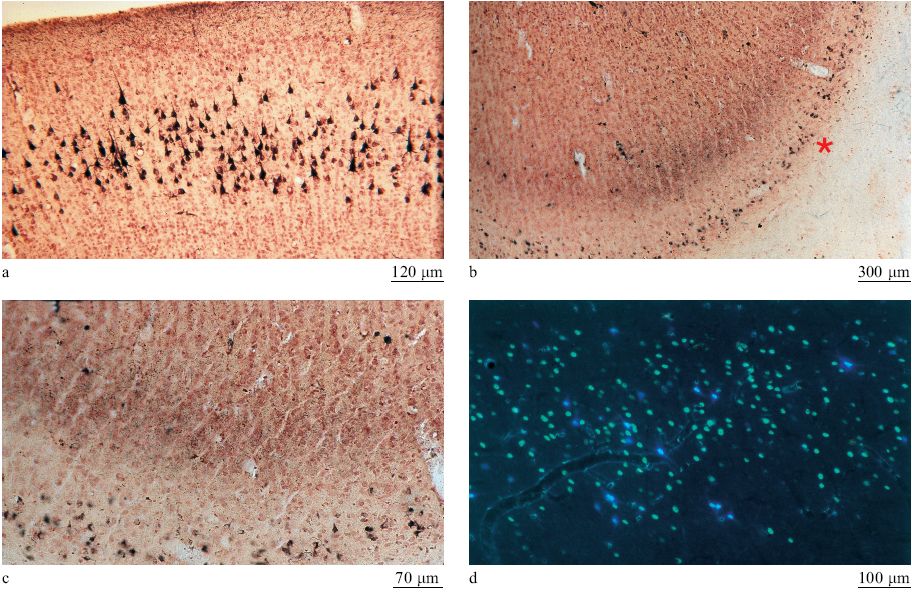
Le aree corti cali comunicano tramite connessioni estrinseche a lungo raggio (quelle che escono dai confini di una certa area) ma hanno anche una fitta rete di connessioni intrinseche (locali, all'interno di un'area) a breve raggio. Tutte le aree ricevono connessioni da molteplici sorgenti. La corteccia è composta di numerose aree che sono distinguibili sulla base di vari criteri. Una classificazione, stabilita nel 19° secolo ma ancora importante, distingue le aree primarie da quelle non primarie. La aree primarie sono strettamente legate ai recettori sensoriali periferici (la retina, la coclea o la pelle) attraverso nuclei sensoriali specifici nel talamo. È probabile che tali aree siano composte di tipi cellulari specializzati, con piccoli campi recettivi e proprietà di risposta relativamente specifiche (fig. 1). Le aree primarie sono organizzate sulla base di mappe topo grafiche riconoscibili (della retina, della coclea o della pelle nel caso di cortecce sensoriali). Una lesione localizzata in una di queste aree porta a deficit defrnibili spazialmente e specifici per una certa modalità sensoriale: per esempio, una lesione della corteccia visiva porta a scotomi nel campo visivo, esattamente come nel caso di danneggiamento della retina o del nervo ottico. Le aree primarie sono facilmente identificabili anche se, a proposito di alcuni sistemi, ancora si discute se più di un'area possa essere considerata primaria. Anche la corteccia non primaria presenta caratteristiche strutturali e fisiologiche riconoscibili, ma queste aree in genere sono più difficili da identificare. Le mappe tendono a essere meno ordinate rispetto al mondo esterno e potrebbero essere organizzate con criteri diversi da quelli topo grafici. La aree non primarie possono essere dominate da una o da più modalità sensoriali (possono essere, cioè, aree unimodali o multimodali). Le aree sensoriali intermedie sono collegate alle aree primarie direttamente o indirettamente, mediante un piccolo numero di connessioni sinaptiche. Le aree superiori stabiliscono connessioni più indirette con le aree primarie. l neuroni in queste aree tendono ad avere proprietà di risposta più astratte, come quelle che potrebbero riguardare, in alcune aree parietali, i contesti spaziali o, in alcune regioni della corteccia frontale e temporale, le funzioni legate alla memoria. Il complesso di queste aree prende anche il nome di corteccia associativa. In passato, si pensava che queste aree fossero importanti per l'integrazione (o l'associazione) di diversi stimoli e concetti.
Struttura cellulare
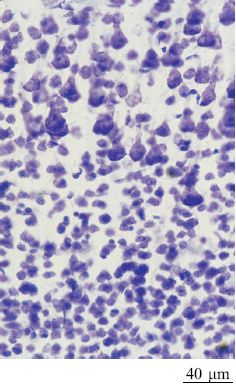
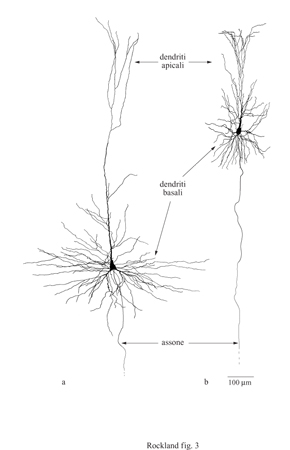
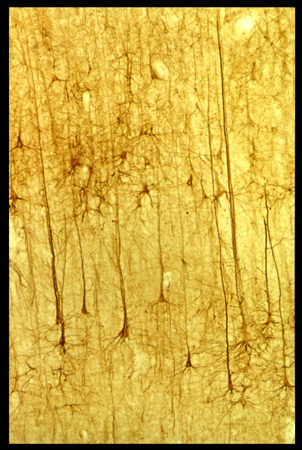
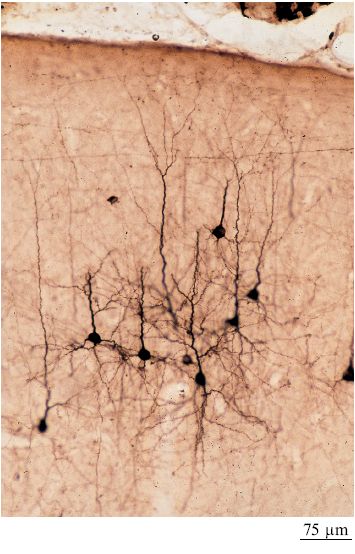
Le aree corticali sono composte principalmente di cellule piramidali (DeFelipe e Fariñas, 1992) che sono tutte eccitatorie, e la maggior parte di esse proietta connessioni estrinseche (attraverso i processi assonali) verso altre strutture corticali o sottocorticali (fig. 2). l neuroni piramidali sembrano piuttosto simili tra loro, ma a un esame più accurato potrebbero essere divisi in sottocategorie sulla base delle connessioni, della morfologia dendritica e assonica, delle proprietà bio fisiche di emissione di impulsi e della distribuzione dei vari marcatori molecolari (figg. 3, 4).
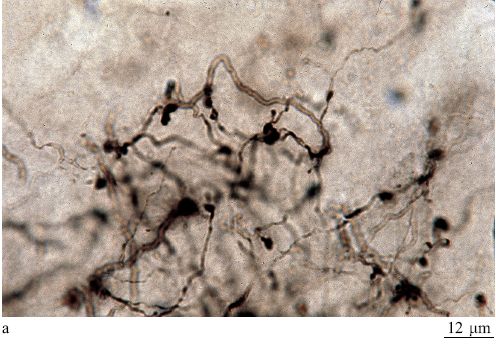
In ogni area, circa il 20% dei neuroni è impegnato in circuiti locali; di qui il nome di interneuroni (White e Keller, 1989; DeFelipe e Fariñas, 1992; Douglas e Martin, 1998). Tali neuroni sono principalmente inibitori e si dividono in un numero compreso tra 6 e 12 di sottocategorie, ognuna caratterizzata da una morfologia dendritica o assonica tipica, da un certo tipo di neuroni a cui si connette preferenzialmente e da un contenuto peptidico distintivo (fig. 5). Gli interneuroni interagiscono con le popolazioni di cellule piramidali con modalità che stiamo appena cominciando a capire. l singoli neuroni potrebbero avere delle proprietà notevolmente specializzate. Le registrazioni elettro fisiologiche da singoli neuroni hanno mostrato un alto livello di specializzazione nelle risposte di queste cellule per suoni, colori e persino per entità complesse come le facce. Tuttavia, nel cervello dei mammiferi si pensa in genere che la funzione corticale dipenda dagli aggregati neuronali o dalle reti, e non da singoli neuroni. Gli aggregati sono organizzati in strati (larghe strisce di circa 0,2 mm, orientate orizzontalmente, parallele alla superficie della corteccia) e colonne (strutture di diversa composizione e misura, orientate verticalmente, ortogonali alla superficie della corteccia).
La struttura a strati
Per convenzione si suppone che le aree corticali abbiano 6 strati, composti da diverse proporzioni di neuroni piccoli, medi e grandi che variano da area ad area. Gli strati hanno anche un diverso livello di connettività: gli strati l e IV ricevono solo connessioni dall'esterno, mentre gli altri contengono neuroni che proiettano le loro fibre in maniera ordinata verso vari bersagli estrinseci. Eccetto che per gli strati l e IV, tutti gli strati hanno connessioni verso l'interno e verso l'esterno. Ogni strato presenta un microambiente distintivo in termini di connessioni anatomiche e di concentrazione di recettori e altre molecole. Gli effetti causati da sostanze come le neurotrofine dipendono dallo strato. Tuttavia è importante ricordare che gli strati non sono delle strutture rigide e chiaramente separabili (come pezzi di legno compensato o come una torta stratificata).l dendriti quasi sempre si estendono negli strati che non contengono il soma da cui si dipartono, e la maggior parte dei neuroni piramidali ha un sistema elaborato di connessioni collaterali dell' assone che si ramificano a partire dall' assone principale (fig. 6). Questo permette molte combinazioni di interazioni intrinseche (all'interno della stessa area) inter- e intralaminari.
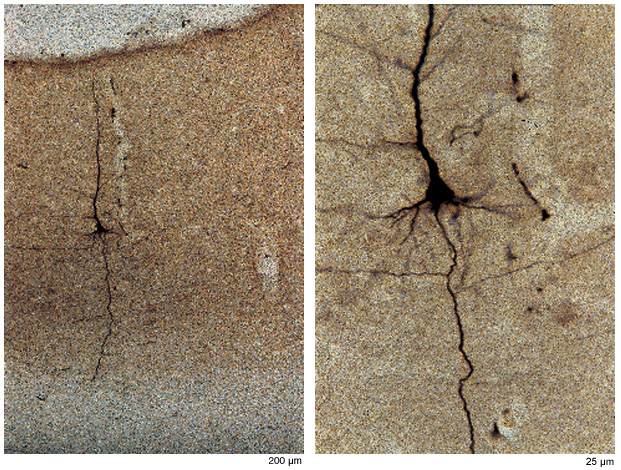
Le connessioni collaterali intrinseche, dette orizzontali perché si irradiano dal soma più o meno ortogonalmente allo strato, mostrano una distribuzione irregolare (Gilbert, 1992). Il sistema delle connessioni collaterali orizzontali è stato studiato in modo estensivo nella corteccia visiva primaria, dove i dati sperimentali suggeriscono che questo sistema potrebbe essere legato alla risposta preferenziale di singole cellule a linee di una certa orientazione. Almeno in alcune specie, le connessioni collaterali dei singoli neuroni si allungano simmetricamente nella corteccia lungo la stessa direzione delle linee per le quali si ha la risposta preferenziale del neurone. Inoltre, i gruppi di cellule con preferenze simili tendono a essere più interconnessi dei gruppi che hanno preferenze opposte, posto che la direzione viene definita in relazione all'asse verticale del campo visivo che si trova lungo il bordo della corteccia visiva primaria. Quindi i campi terminali che sono allineati parallelamente al bordo corticale possono essere considerati come orientati lungo l'asse del campo verticale. Si pensa che questo tipo di struttura giochi un ruolo importante nell' affrnare la precisione della curva di risposta all' orientazione al livello di campi recettivi (figg. 7, 8).
La funzione delle connessioni collaterali non è ancora stata chiarita, sebbene questo sistema sia l'elemento di base dell' architettura corticale. Dal momento che il sistema di organizzazione orizzontale esiste in altre aree, oltre che nella corteccia visiva, si può ragionevolmente dedurre che il sistema abbia una qualche funzione comune e astratta, o che sia legato a diverse funzioni in aree differenti. Un'ipotesi è che il sistema orizzontale sia coinvolto in effetti di contorno, chiamati anche effetti contestuali (Gilbert, 1992), come i mutamenti nell'attività di scarica di un neurone in risposta a quanto sta accadendo nelle regioni vicine dello spazio. Questa ipotesi è confermata dal fatto che le connessioni collaterali orizzontali si estendono per almeno 2-6 mm, ben oltre la dimensione dei classici campi recettivi sensoriali.
È stato anche ipotizzato che le connessioni orizzontali possano servire a propagare pattern di attività corticale. Questi pattern vengono studiati in esperimenti che usano dei coloranti molto sensibili al voltaggio in combinazione con la tecnica di visualizzazione ottica (v. anche il saggio di L. Maffei e N. Berardi, in questo volume). In questo modo è possibile visualizzare i cambiamenti della depolarizzazione che risultano dall'attività elettrica di milioni di neuroni mentre essi rispondono a diverse caratteristiche spaziali o temporali di uno stimolo esterno. Questi esperimenti mostrano che le risposte corticali a stimoli poco estesi si diffondono rapidamente dal luogo iniziale (alla velocità di 100÷250 μm/ms), fino a coprire un'area almeno dieci volte più estesa (Grinvald et al., 1994). Ciò suggerisce la metafora della corteccia come moire o come palinsesto di pattern di attività correnti, entranti e preesistenti.
La struttura a colonne
Le aree corti cali hanno anche una struttura verticale. Gli aggregati neuronali chiamati colonne o moduli hanno in comune connessioni anatomiche e proprietà di risposta fisiologica (fig. 9). Le colonne, come gli strati, sono entità complesse. In alcune aree i neuroni stessi sono organizzati in sottili colonne o in catene, forse legate a gruppi di neuroni con sviluppo comune (doni) ed è stato supposto che i dendriti apicali di questi neuroni formino delle minicolonne di circa 50 μm di diametro, importanti dal punto di vista funzionale (Peters e Sethares, 1991). Questi dati sperimentali sostengono l'interpretazione che le colonne cellulari potrebbero essere le unità di base o i mattoni costitutivi elementari dei processi corti cali globali. Tuttavia non è necessario che le colonne siano delle unità ripetute rigidamente (come l'ommatidio dell'occhio degli insetti). Altri dati suggeriscono che le colonne sono in effetti molto diversificate e che dunque parecchie strutture diverse vengono sussunte sotto il nome generico di colonne.
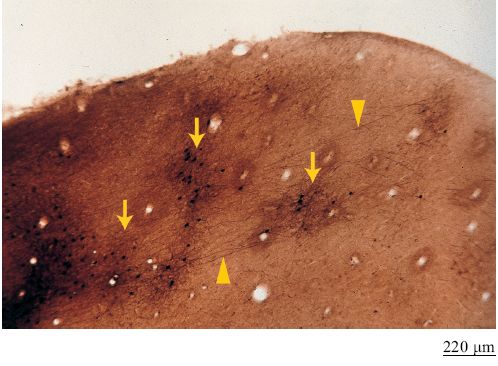
Nelle aree sensoriali primarie le colonne corrispondono esattamente ad alcuni aspetti dell'attività sensoriale periferica, e in queste aree le colonne sembrano basarsi su connessioni talamo-corticali. Per i roditori, per esempio, che sono dotati di vibrisse, questo sistema di sensori fornisce uno strumento importante per stabilire la struttura dell'ambiente, e perciò in questi animali la corteccia primaria somatosensoriale contiene una mappa o una rappresentazione dettagliata dell'area del muso (Woolsey e Van der Loos, 1970). La disposizione spaziale delle singole vibrisse viene letteralmente replicata nel tessuto corticale, grazie alla convergenza di fibre nervose da ogni vibrissa a centinaia di cellule dello strato IV. Queste colonne vennero inizialmente individuate tramite semplice colorazioni delle cellule negli anni Quaranta. Poiché questi raggruppamenti cellulari presentano sotto strutture istologicamente distinte in pareti e cavità, ovvero fibre talamiche di input circondate da pareti di cellule, sono anche conosciuti come 'barili'.
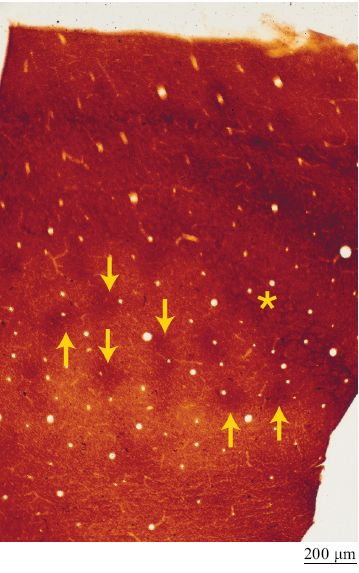
Nei primati non si riscontra un comportamento analogo al movimento delle vibrisse nei roditori, ma un livello di specializzazione equivalente si è evoluto per la vista. Si ritiene che la corteccia visiva primaria dei primati abbia almeno due sistemi colonnari (Rubel e Wiesel, 1977), corrispondenti alla dominanza oculare (competizione tra occhio destro e sinistro) e all' orientazione degli stimoli percepiti (come i singoli neuroni rispondono a linee orientate con diversi angoli). Il sistema di dominanza oculare, come il campo recettivo dei 'barili' nei roditori, corrisponde a insiemi di fibre talamiche di input che convergono su aggregati di neuroni dello strato IV (gli insiemi sono larghi circa 0,4 mm). Questo sistema può essere visualizzato in proiezioni piatte della corteccia usando tecniche opportune. Un altro sistema, non strettamente colonnare, consiste in chiazze di sfere irregolari centrate sulle colonne di dominanza oculare (fig. 10). Queste chiazze corrispondono a regioni con elevata attività metabolica e possono essere visualizzate mediante tecniche di immunoistochimica sensibili a enzirni come la citocromoossidasi altri marcatori. Il sistema per la selettività all' orientazione comprende due schemi di rappresentazione: piccoli segmenti lineari o tavolette che in sezione appaiono come colonne, e strutture chiamate singolarità, dove convergono tutte le orientazioni preferenziali (Blasdel, 1992). l due sistemi riguardano rispettivamente le informazioni visive precise su punti locali e le informazioni più approssimative su tutte le orientazioni. Il substrato del sistema per la selettività all' orientazione è una combinazione di connessioni cortico-corticali e talamo-corticali. Sono disponibili anche dati sperimentali che mostrano l'esistenza di colonne in aree di livello superiore. Nella corteccia inferotemporale, una regione visiva di alto livello gerarchico, sono presenti dei raggruppamenti di cellule che rispondono ad alcune caratteristiche visive degli oggetti (Tanaka, 1997). Tuttavia, al contrario della mappatura topo grafica caratteristica dei sistemi intermedi, si pensa che la mappatura realizzata in quest' area si basi su di uno spazio delle caratteristiche (fig. 11). Le caratteristiche possono essere semplici, come quelle a cui rispondono i neuroni della corteccia visiva primaria (per esempio, l'orientazione, la dimensione, il colore o la struttura del motivo grafico che ricopre la superficie) oppure, come avviene tipicamente in quest'area, sono combinazioni di forme più complesse.
Le colonne sono spesso considerate come strutture solide, come assi da carpentiere o chip di un computer, ma, come nel caso degli strati, questa è soltanto una semplificazione. Per ogni gruppo locale di colonne si possono creare delle ulteriori suddivisioni selettive, attraverso complesse interazioni dipendenti dal tempo tra le varie parti dello spazio corticale. Ciò viene illustrato, per esempio, dai risultati degli esperimenti svolti sui pattern di attività elettrica nella corteccia somatosensoriale dei roditori in risposta a diverse configurazioni di deflessione delle vibrisse. Tra l'altro si osserva che il campo di attività è non lineare, nel senso che la deflessione alternata di due vibrisse vicine provoca un'intensità di risposta minore della somma delle intensità corrispondenti alla deflessione di ogni singola vibrissa (Kleinfeld e Delaney, 1996).
È possibile che delle unità di così piccole dimensioni come le colonne possano influenzare il comportamento di un organismo senziente? In un interessante approccio sperimentale a questo problema è stata confrontata la sensibilità psicofisica di una scimmia addestrata a osservare e a rispondere a stimoli visivi con la sensibilità di un singolo neurone in una delle aree visive non primarie (Britten et al., 1992; Salzman et al., 1992). Si è così giunti alla conclusione che la risposta di un tipico neurone potrebbe fornire una spiegazione accurata delle prestazioni psicofisiche della scimmia. Quando le risposte neuronali venivano manipolate tramite microstimolazione della corteccia, le discriminazioni psicofisiche dell' animale potevano essere alterate in modo predicibile. In questo e in altri esperimenti simili è stato stimato che il numero minimo di neuroni necessari a influenzare le prestazioni comportamentali è relativamente piccolo (circa 25).
Connessioni corticali
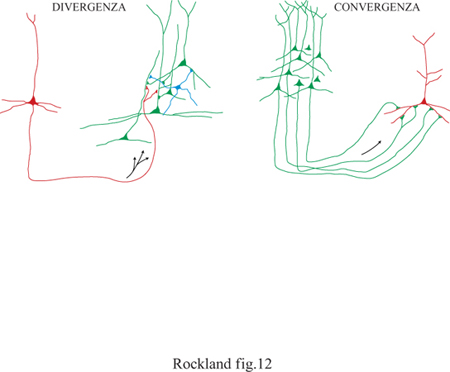
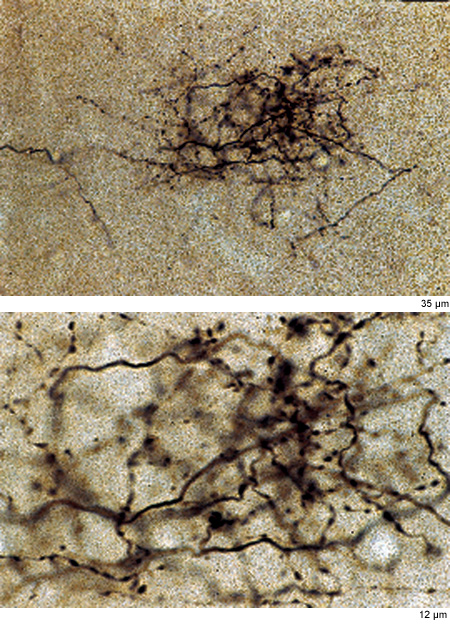
l neuroni piramidali possiedono lunghi assoni che vanno a connettersi a bersagli lontani millimetri o centimetri. A loro volta, i singoli neuroni sono immersi in una rete complessa di connessioni estrinseche. Le connessioni sono caratterizzate da convergenza e divergenza (fig. 12). l tipici neuroni piramidali divergono verso parecchie centinaia o addirittura migliaia di altri neuroni (fig. 13). A sua volta, ogni neurone riceve più di 6000 sinapsi (input convergenti) che provengono da diverse sorgenti estrinseche, sebbene la maggior parte (circa 1'80%) venga da neuroni vicini piramidali o non piramidali (White, 1989; DeFelipe e Fariñas, 1992; Rockland, 1997; Douglas e Martin, 1998).
La convergenza e la divergenza delle connessioni sono fondamentali per le operazioni neuronali, e rappresentano una sorta di sintassi connettiva. È disponibile una quantità considerevole di dati sull'identità e l'organizzazione di sistemi connettivi individuali, sebbene per la maggior parte dei sistemi la ricerca sia ancora impegnata nel decifrare i principi e i fatti più importanti. La questione di vitale importanza riguardante i principi che regolano tali interazioni è quella più difficile e ancora irrisolta, ma è stata affrontata in diversi modi.
Identificazione del tipo di connessione
L'insieme di base dei vari tipi di connessioni è noto per la maggior parte delle aree corti cali. Questo dato è stato ottenuto principalmente mediante tecniche istologiche, iniettando piccolissime quantità di sostanze traccianti nel liquido extracellulare nei cervelli di soggetti vivi. Queste sostanze vengono trasportate lungo l'assone (in direzione anterograda e retrograda) a partire dalla zona di iniezione per via dei normali processi fisiologici di trasporto assonale, e vengono visualizzate con speciali procedure di marcatura. In questo modo è possibile mostrare che due aree sono connesse tra loro e, più precisamente, è possibile identificare i neuroni da cui parte la connessione e stabilire come gli assoni terminano nella regione bersaglio.
Tutte le aree ricevono numerose connessioni estrinseche. Per esempio, la corteccia visiva primaria è strettamente connessa con il talamo visivo, ma riceve anche input diretti da numerose altre strutture corticali e sottocorticali. Le connessioni verso le aree non primarie sono persino più numerose. Tuttavia, la maggior parte degli studi riguarda le fibre che viaggiano dal talamo alla corteccia (connessioni talamo-corticali) e quelle che legano tra loro diverse aree cortic ali (connessioni cortico-corticali). Il discorso che segue sarà incentrato su questi due sistemi come esempi rappresentativi.
Le connessioni talamo-corticali. - Il talamo è un'importante e cospicua struttura sottocorticale che comprende dei sottonuclei reciprocamente connessi a singole aree corticali. I nuclei sensoriali specifici ricevono connessioni dagli organi sensoriali periferici e a loro volta proiettano fibre verso le aree sensoriali primarie della corteccia. Talvolta conviene parlare di relé talamici dalla periferia al mondo esterno, sebbene l'organizzazione di queste connessioni sia più complicata di semplici relé.
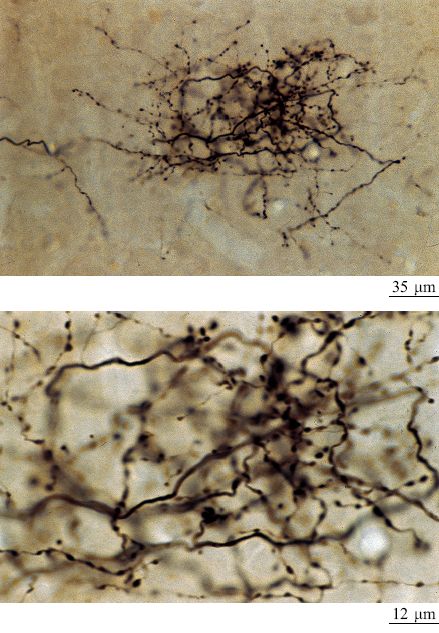
Le connessioni talamo-corticali (CTC) sono la principale sorgente di input estrinseci alle aree sensoriali primarie e sono essenziali per le risposte dei neuroni specifiche per ogni modalità sensoriale. Questo significa che l'eliminazione degli input talamici, ottenuta tramite lesioni, raffreddamento o disattivazione farmacologica del talamo, provoca la perdita della sensibilità (risposta dei neuroni) visiva, uditiva o somatosensoriale delle aree primarie, come è confermato dalle registrazioni elettrofisiologiche. Si pensa che le CTC siano importanti per stabilire le mappe topo grafiche che caratterizzano le aree primarie. Le CTC possono essere suddivise in diversi tipi, corrispondenti a varie proprietà sensoriali specifiche, diversi strati di terminazione nella corteccia e diverse morfologie di arborizzazione (Blasdel e Lund, 1983). Per esempio, nella corteccia visiva alcuni assoni delle CTC sono coinvolti nella percezione del colore e della forma e presentano arborizzazioni terminali relativamente piccole (fig. 14). Altri assoni invece sono coinvolti nella percezione del movimento degli stimoli e presentano arborizzazioni di dimensioni doppie. Questi due tipi di fibre possono essere pensati come estensioni dei due sistemi retinici di coni e bastoncelli. In questo contesto le piccole arborizzazioni possono determinare risoluzioni più elevate, mentre le arborizzazioni più estese, che presumibilmente coinvolgono un numero maggiore di neuroni, possono portare a una sensibilità più alta.
Le CTC costituiscono anche uno degli input principali per le aree non primarie. Queste connessioni partono da nuclei non primari o di associazione all'interno del tal amo e sono state meno studiate, sia anatomicamente che fisiologicamente, di quelle che si originano nei nuclei sensoriali specifici. Tali fibre potrebbero essere coinvolte nella formazione delle mappature topo grafiche, ma potrebbero anche riguardare processi più astratti.
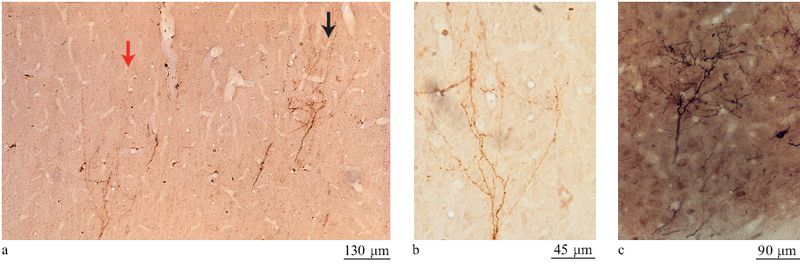
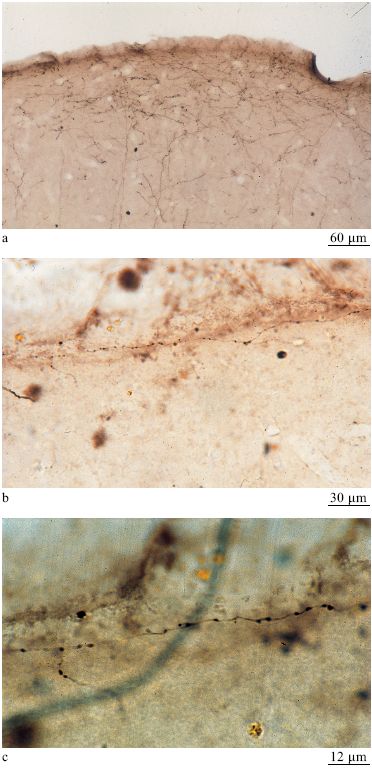
Le connessioni cortico-corticali. - Queste connessioni formano una rete densa e ordinata di connessioni dirette e indirette tra aree corticali. l singoli neuroni di un'area spesso sono connessi a un'altra sola area, sebbene una piccola parte di essi si connetta a più di un'area corticale diversa o anche a strutture corticali e sottocorticali attraverso ramificazioni collaterali dell'assone principale. Gruppi di neuroni si connettono a diverse strutture bersaglio in modo che una singola area proietti le proprie connessioni in modo divergente verso bersagli multipli. Le connessioni cortico-corticali sono divise in diverse sottopopolazioni: connessioni feedforward (con propagazione in avanti), connessioni ricorrenti e connessioni laterali (fig. 15) (Felleman e Van Essen, 1991; Salin e Bullier, 1995; Rockland, 1997). La scelta di questi nomi deriva principalmente da criteri anatomici, cioè dai pattem laminari e dalle configurazioni di arborizzazione, e relativamente poco si sa sulla fisiologia corrispondente. In termini di strati, le connessioni feedforward si dipartono dai neuroni dello strato III e terminano principalmente nello strato IV (fig. 16), mentre le connessioni ricorrenti partono soprattutto dai neuroni dello strato VI e terminano principalmente nello strato I (fig. 17). In termini di campi terminali, le connessioni feedforward hanno una o più arborizzazioni di ridotte dimensioni, in genere di circa 0,2 mm di diametro, mentre le connessioni ricorrenti tendono ad avere campi di terminazione ristretti, sul tipo di quelli dei bastoncelli, che si estendono su zone di 1÷2 mm. Le connessioni laterali si originano principalmente da neuroni degli strati III e V e tendono ad avere terminazioni a disposizione colonnare che coinvolgono diversi strati. Le colonne di terminazione, che tipicamente hanno un diametro di circa 0,25÷0,50 mm, consistono in centinaia di assoni convergenti; l'estensione e la distribuzione dell'arborizzazione non sono ancora conosciute al livello di assoni individuali.

Si presuppone che tutti questi sistemi siano eccitatori, dal momento che partono da neuroni piramidali. Queste connessioni terminano principalmente su altri neuroni piramidali, ma probabilmente circa il 20% dei loro contatti sono su interneuroni inibitori. Le arborizzazioni cortico-corticali tendono ad avere una densità di terminazioni più bassa di quelle delle CTC, e le singole terminazioni (possibili contatti sinaptici) sono più irregolari (fig. 18). Il significato funzionale di queste differenze non è ancora noto. Le connessioni laterali sono caratteristiche delle connessioni tra aree di alto livello gerarchico. Le connessioni feedforward e quelle ricorrenti sono più specifiche per le cortecce sensoriali primarie e spesso sono bidirezionali tra coppie di aree, sebbene non necessariamente tra gli stessi due neuroni o gruppi di neuroni. Le denominazioni feedforward e ricorrente sono un modo sintetico di ricordare che, da un lato, almeno per le cortecce sensoriali, l'attività parte da stimoli periferici prodotti dal mondo esterno e poi procede attraverso alcune stazioni neuronali di complessità crescente (in avanti); dall'altro, le connessioni bidirezionali o 'di ritorno' esercitano una qualche influenza, modulatoria o di rinforzo, sulle connessioni feedforward (sono ricorrenti). In realtà, questi due sistemi fanno parte di un'intricata matassa di connessioni ed è improbabile che funzionino l'uno indipendentemente dall'altro. La direzionalità della connessione può essere ingannevole in quanto gli stimoli esterni non innescano un'attività neuronale partendo da uno stato di riposo, ma si sovrappongono a un'attività già presente. Questa attività può essere prodotta sia da stati interni (come nell'immaginare di vedere una scena o un oggetto) che da stimoli esterni (come nella percezione di stimoli reali), e in alcune condizioni l'influenza delle connessioni ricorrenti può essere predominante.
Interazioni tramite connessioni
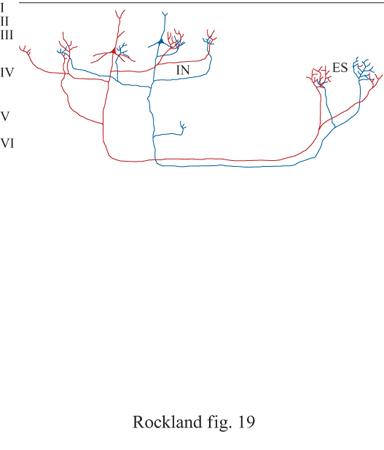
Le connessioni vengono spesso schematizzate, per amore di semplificazione, come piccoli tubi o come percorsi tra diverse regioni. Questa rappresentazione coglie graficamente la natura fisica delle connessioni, se queste vengono intese come fasci di assoni più o meno definiti. Ha invece meno successo nell'esprimere i complessi aspetti temporali e dinamici, come quelli che potrebbero essere rivelati da un'analisi di correlazione incrociata tra diversi siti. Questa schematizzazione non tiene nemmeno conto del fatto che le connessioni estrinseche della maggior parte dei neuroni sono essenzialmente rami di connessioni collaterali intrinseche, e che devono esistere consistenti interazioni tra queste due parti del sistema (fig. 19). Il semplice groviglio delle connessioni e le loro interrelazioni, oltre alla loro complessità funzionale, rendono non facile il tentativo di studiare tali interazioni. Nonostante queste difficoltà, sono stati sviluppati diversi eleganti protocolli sperimentali per lo studio delle interazioni in vari preparati, in vitro, negli animali o negli esseri umani.
Esperimenti in vitro. - Le colture di tessuti o di organi sono di limitata utilità ai fini dello studio delle interazioni fra aree o centri cerebrali, a causa delle grandi distanze che separano la struttura sorgente e la struttura bersaglio. Tuttavia in alcuni casi è possibile includere grosse porzioni di regioni interconnesse in una fettina isolata di tessuto cerebrale. Ciò permette di eseguire manipolazioni sperimentali come la stimolazione elettrica focale. La propagazione dell'attività neuronale che viene prodotta può essere visualizzata usando coloranti sensibili al voltaggio in combinazione con tecniche di visualizzazione ottica in tempo reale. È possibile per esempio dissezionare la corteccia entorinale insieme all'ippocampo (lijima et al., 1996). La corteccia entorinale è un'area che fa parte del lobo temporale mediale del cervello, riceve connessioni da diverse altre aree ed è connessa all'ippocampo. La stimolazione elettrica della corteccia entorinale in queste condizioni porta a pattern di attività neuronale che sono interpretati come l'interazione di almeno due circuiti: uno che costituisce un circuito riverberante all'interno della stessa corteccia entorinale, e l'altro che risulta essere composto da connessioni estrinseche che vanno dalla corteccia entorinale all'ippocampo.
Sono stati osservati effetti di plasticità nelle connessioni orizzontali delle fettine di tessuto (Hirsch e Gilbert, 1991). La stimolazione elettrica delle connessioni intrinseche produce risposte eccitatorie o inibitorie di diversa intensità. Per stimoli di bassa intensità l'effetto è quasi esclusivamente eccitatorio, ma all'aumentare della tensione elettrica dello stimolo l'inibizione tende progressivamente a predominare. Così, l'effetto dell' attivazione delle connessioni orizzontali non è costante, ma può cambiare sulla base del numero di fibre coinvolte e dello stato di attivazione delle cellule postsinaptiche. Effetti di plasticità analoghi si verificano con molta probabilità in altri sistemi, ma risulta tecnicamente difficile riuscire a dimostrarlo.
Esperimenti condotti su animali. - Il metodo più comune per indagare sulle interazioni fra aree cerebrali è quello adottato negli esperimenti di disattivazione, che sono progettati per indagare sull'azione esercitata da singoli sistemi sulle loro aree bersaglio. Gli effetti di tale trattamento vengono studiati a livello di proprietà neuronali mediante registrazioni elettro fisiologiche, oppure osservando i cambiamenti nel comportamento globale dell'animale in condizioni di test accuratamente controllate. La disattivazione progressiva può talvolta servire a sondare le interazioni fra due sorgenti di input.
Si potrebbe pensare che le connessioni corticali feedforward abbiano per le aree bersaglio un ruolo analogo a quello che le connessioni talamiche hanno per la corteccia primaria, ovvero un'influenza essenziale o 'di controllo'. Tuttavia la disattivazione della corteccia visiva primaria (area VI) elimina la capacità di produrre risposte visive in alcune, ma non tutte, le aree visive non primarie che ricevono connessioni da essa (Bullier et al., 1994; Salin e Bullier, 1995). Per queste aree, o gli input che provengono dalla corteccia primaria non sono essenziali (cioè hanno un ruolo secondario rispetto ad altri input), oppure la loro mancanza può essere compensata da connessioni con altre aree con le quali normalmente interagiscono. Così un'area visiva di livello gerarchico abbastanza elevato, come l'area mediotemporale (MT) o V5, può mantenere inalterate le sue proprietà di risposta anche dopo la disattivazione dell'area VI, ma i suoi neuroni mostreranno deficit se si combina la disattivazione di VI con quella degli input talamici (attraverso il collicolo superiore e il pulvinar).
Anche le connessioni ricorrenti sono state studiate mediante esperimenti di disattivazione. Disattivando le connessioni che vengono da V2 (un'area del sistema visivo strettamente connessa a VI) si producono effetti definiti sulle proprietà dei campi recettivi di VI (Bullier et al., 1996). La disattivazione porta all'aumento della risposta della periferia e alla diminuzione della risposta del centro dei campi recettivi. Presumibilmente questo effetto fa diminuire la selettività per piccoli stimoli dei neuroni di VI. In un altro esperimento, condotto su scimmie, le connessioni ricorrenti che arrivano alla corteccia inferotemporale dalla corteccia entorinale e peririnale sono state interrotte provocando delle lesioni (Higuchi e Miyashita, 1996). Gli effetti sono stati studiati mediante registrazioni elettrofisiologiche di risposte neuronali singole mentre le scimmie eseguivano un compito che richiedeva l'associazione visiva di due insiemi di figure. Dopo il danneggiamento delle connessioni ricorrenti, la capacità dei neuroni della corteccia inferotemporale di rispondere a certe immagini rimaneva inalterata, mentre veniva persa la capacità di codificare le associazioni tra coppie di figure.
Recentemente, sono state sviluppate nuove tecniche per impiantare degli elettrodi molto sottili in diversi punti del cervello, grazie alle quali è possibile controllare l'attività neuronale negli animali durante l'esecuzione di comportamenti naturali (Nicolelis et al., 1995). Questa tecnica mostra in modo chiaro l'esistenza di complessi pattern spazio-temporali di attivazione in vari luoghi della corteccia e del talamo. Sarà tuttavia necessaria un'analisi ulteriore per determinare precisamente le interazioni.
Un'altra nuova e promettente tecnologia viene offerta dalle manipolazioni genetiche, di solito effettuate sui roditori (Wilson e Tonegawa, 1997). Per esempio, in un ceppo di topi mutanti recentemente scoperto gli assoni talamo-corticali non riescono a segregarsi negli usuali pattern a chiazze che corrispondono alle vibrisse del muso dell' animale (Welker et al., 1996). Nonostante questo difetto anatomico, si può dimostrare funzionalmente l'esistenza nella corteccia di una mappa topo grafica della zona delle vibrisse. Alcune anomalie corticali vengono rivelate dalle registrazioni da singole cellule, che mostrano una degradazione non tanto nella topografia quanto piuttosto nell'organizzazione temporale del centro e della periferia dei campi recettivi. Questo risultato suggerisce sorprendentemente che per l'analisi spazio-temporale è necessario avere una precisa organizzazione delle terminazioni talamo-corticali, ma anche che questa organizzazione è meno essenziale per l'ordinamento topografico o che può almeno essere compensata in altri modi.
Esperimenti condotti su esseri umani. - La ricerca sugli esseri umani può riguardare una grande varietà di comportamenti diversi, ma allo stesso tempo risulta più difficile stabilirne i fondamenti neurobiologici, perché la maggior parte delle usuali tecniche è troppo invasiva per poter essere usata su esseri umani. Un modo di aggirare questo problema è stato quello di analizzare attentamente i deficit che conseguono a un danno cerebrale localizzato, una sorta di esperimento di disattivazione fatto dalla natura. Il metodo di analisi delle lesioni può fornire uno strumento sensibile per studiare le reti nervose negli esseri umani (Damasio, 1994). Tanto per fornire un esempio tra i molti possibili, è noto che lesioni bilaterali della regione occipito-temporale mediale portano a una condizione nota come prosopoagnosia. L'aspetto più drammatico del deficit è l'incapacità del paziente di riconoscere le facce. Tuttavia, dal momento che la percezione visiva e la memoria rimangono intatte, non è appropriato parlare della perdita di un'area che memorizza 'fotograficamente' facce (una sorta di 'area delle facce'). Questo disturbo si può meglio caratterizzare come una perdita del senso di familiarità, o come un'evocazione contestuale difettosa per certi stimoli, come quelli che appartengono a una categoria visivamente ambigua: per esempio, diversi tipi di automobili o di facce (Damasio et al., 1982). Il processo che fa percepire come familiare uno stimolo, e che è coinvolto nella prosopoagnosia, può essere analizzato come un processo a tre stadi, durante il quale uno stimolo viene dapprima percepito, poi associato correttamente a uno stimolo memorizzato e infine accompagnato da un'evocazione di informazioni contestuali (v. oltre, a proposito delle zone di convergenza). Si può dunque inferire che il processo coinvolga diversi insiemi di connessioni (callo sali, talamocorticali e cortico-corticali) e che la distruzione o l'alterazione di queste interazioni provochi il deficit cognitivo. Una strategia di indagine complementare consiste nello studiare i pattern di attività neuronale negli esseri umani usando tecniche di visualizzazione dell'attività cerebrale in vivo, come la PET e la tMRI. Questi studi confermano il coinvolgimento di vari siti del cervello anche nello svolgimento di compiti semplici e sollevano la questione delle dinamiche sotto stanti.
Elaborazione di modelli formali. - La modellizzazione delle connessioni e delle interazioni tra le aree cerebrali fornisce un altro importante strumento di indagine. Sono stati sviluppati parecchi modelli delle interazioni dipendenti dalle connessioni feedforward (Ullman, 1995), talamo-cortic ali e cortico-corticali (Mumford, 1992). Simulazioni mediante reti neurali molto estese possono tentare di gettare un ponte tra la struttura dei singoli neuroni e sinapsi e la dinamica di grossi insiemi di neuroni che conduce ai comportamenti osservabili (Wray ed Edelman, 1996; Amit e Brunel, 1997). Le simulazioni, finora, hanno dato rilievo allo studio della dinamica collettiva di neuroni semplificati, senza tenere conto delle relazioni tra dinamica e comportamento. Per esempio, tramite le simulazioni sono state studiate le condizioni di stabilità per l'attività spontanea in una rete non strutturata, o la sua risposta a una perturbazione indotta da uno stimolo esterno. Questi modelli diverranno più realistici quando terranno conto dei dati sperimentali e si avvarranno dello sviluppo di nuovi algoritmi.
Architetture su larga scala
Come abbiamo detto in precedenza, i sistemi neuronali sono stati studiati principalmente a un livello analitico. È tuttavia importante procedere rapidamente verso una sintesi teorica coerente. Diversi quadri teorici di riferimento sono stati già proposti per collegare l'organizzazione cognitiva a quella neuronale al livello di sistemi su larga scala.
La struttura gerarchica
Secondo un punto di vista molto accreditato tra gli studiosi, la corteccia è organizzata secondo una struttura gerarchica (Felleman e Van Essen, 1991). Questa opinione è compatibile con la molteplicità delle aree corticali sensoriali e con ciò che sembra essere un'elaborazione progressiva delle proprietà di risposta dei neuroni, a mano a mano che ci si sposta dalle aree primarie a quelle non primarie (superiori). Nella formulazione più semplice di questo principio, una struttura gerarchica implica una progressione seriale dalle aree primarie sensoriali alle aree superiori. Le connessioni talamo-corticali che arrivano alle aree sensoriali primarie sono considerate come il punto di partenza di questo processo o come il livello più basso, mentre negli stadi successivi il segnale viene elaborato mediante connessioni cortico-corticali feedforward e altre connessioni. Le connessioni reciproche feedforward e ricorrenti, organizzate a coppie, costituiscono importanti indizi per stabilire il livello gerarchico di un'area.

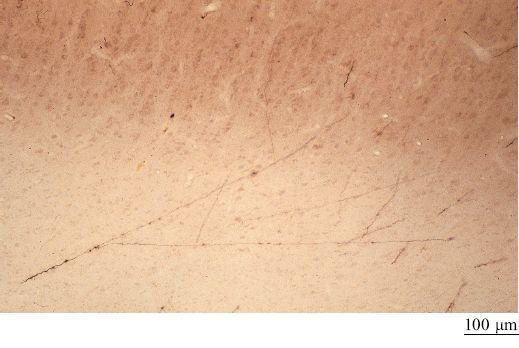
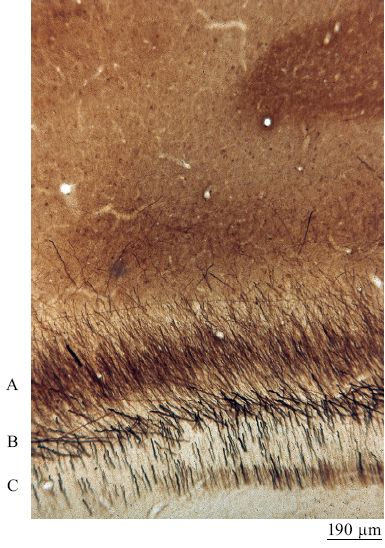
Nonostante una struttura di relazioni così ordinata sia attraente, questa formulazione non tiene conto di parecchi fatti sperimentali osservati (Rockland, 1997). Per esempio, diverse aree, sebbene siano spesso interconnesse reciprocamente tra loro, possono interagire anche in altri modi. Tra questi vi sono gli assoni ramificati, che divergono verso due o più aree separate, e i circuiti chiusi composti da connessioni talamo-corticali e da altre connessioni, sia dirette che indirette (fig. 20). Inoltre, ogni sistema di connessioni non è uniforme. Anatomicamente le connessioni consistono in assoni di diverso calibro, cui corrispondono diverse velocità di conduzione (figg. 21, 22). Quindi gli esperimenti di fisiologia riportano una grande variabilità delle latenze di risposta a uno stimolo all'interno di una stessa area cerebrale e in aree diverse. Questi esperimenti mostrano che in media esiste un ritardo di 10 ms per passare da una stazione all'altra lungo il percorso di aree corticali interconnesse.
Tuttavia, sono necessarie delle precisazioni. Nel sistema visivo corticale, nel momento in cui il 50% dei neuroni di VI ha risposto alla presentazione dello stimolo, ha risposto anche il 43% dei neuroni di V2 (Nowack e Bullier, 1997).
Inoltre, alcuni neuroni nella corteccia inferotemporale possono rispondere a uno stimolo persino prima di alcuni neuroni di VI. Sembra dunque che vi sia una ricchezza di combinazioni di connessioni. Tuttavia le strutture gerarchiche sono compatibili con l'esistenza di processi distribuiti e paralleli, e con diverse modalità di interazione, sia locali che globali. l governi o le aziende vengono spesso citati come esempi di questo principio di organizzazione gerarchica. In effetti, sebbene si pensi che le aree siano le unità organizzative principali della corteccia, risulta evidente che esse si possono raggruppare in famiglie funzionalmente omogenee (Levitt et al., 1997).
Alcuni di questi complessi formati dalla connessione fra più aree hanno in comune dei marcatori molecolari. Per fare soltanto qualche esempio, la proteina LAMP (Limbic-system Associated Membrane Protein, proteina di membrana associata al sistema limbico) viene prodotta da regioni della corteccia che classicamente sono considerate limbi che sia per connessioni che per funzione (cortecce peririnale, cingo lata e prefrontale del ratto). Un'altra proteina, CAT-30l, viene prodotta dalle suddivisioni del sistema visivo primario che sono associate, per connessioni e per funzione, alla percezione visiva del movimento.
La natura problematica dell'organizzazione gerarchica del cervello richiama quello che in un altro contesto è stato chiamato il fenomeno degli strani anelli o delle gerarchie aggrovigliate. Per esempio, riferendosi alle paradossali figure create da M.C. Escher, D.R. Hofstadter (1984) afferma: "Certo si può pensare che ci sia una qualche ambiguità nella nozione di numero di componenti: per esempio, in Salita e discesa non sarebbe possibile individuare quattro livelli (le rampe) piuttosto che quarantacinque (i gradini)?".
La struttura delle zone di convergenza
Al di là dei dettagli concernenti l'organizzazione neuronaIe, si pone il problema di come gli stati e le attività mentali emergano dal substrato neuronale. Come nel 'problema del legame' (binding problem), il punto è chiedersi come esperienze uniformi e integrate scaturiscano da pattern neuronali spazialmente e temporalmente eterogenei che vengono scatenati da particolari stimoli ambientali (Singer, 1994).
Nella struttura delle zone di convergenza dell'organizzazione cerebrale (Damasio, 1989; 1994; Damasio e Damasio, 1994) è importante distinguere tra rappresentazioni topograficamente organizzate, associate alle cortecce primarie sensoriali e motorie, e pattern neuronali disposizionali, associati ad aree corti cali di livello superiore e a nuclei sottocorticali. Questi ultimi consistono in pattern potenziali di attività nervosa in piccoli gruppi di neuroni (le zone di convergenza); tali zone di convergenza organizzano o impongono un ordine nei pattern neuronali prodotti in altre aree con cui esse sono strettamente connesse (fig. 24). Queste connessioni hanno a che vedere con regole relazionali, piuttosto che con il contenuto in informazione.
Nelle parole di A.R. Damasio (1994): "Quel che le rappresentazioni disposizionali tengono immagazzinato nella loro minuscola comune di sinapsi non è 'una figura' in sé, ma un mezzo per ricostruire una figura. Il fatto di avere una rappresentazione disposizionale del volto di zia Margherita non significa che la rappresentazione contenga quel volto, ma piuttosto gli schemi di scarica che innescano la ricostruzione istantanea di una rappresentazione approssimativa proprio del volto di zia Margherita nelle cortecce visive di ordine inferiore" .
In questo contesto, l'organizzazione temporale gioca un ruolo importante, e le immagini mentali si spiegano come costrutti momentanei e non come riproduzioni immagazzinate. Le immagini mentali esplicite emergono dall'attivazione transitoria e sincronica di pattern di scarica neuronale nelle stesse cortecce sensoriali dove, precedentemente, venivano prodotte le rappresentazioni percettive. Le rappresentazioni topografiche sono così distinte dalle disposizioni, ma si assume che entrambe agiscano necessariamente di concerto.
Questa proposta teorica è stata in parte elaborata a partire dall' analisi sistematica delle conseguenze comportamentali di lesioni selettive, ed essa assume come presupposto che esista una corrispondenza consistente tra certi sistemi neuronali e certi tipi di conoscenza. La relazione tra gerarchie (o livelli di complessità) di conoscenza e gerarchie di aree corticali è considerata ragionevole, se non proprio esatta. Per esempio, l'accesso a una conoscenza concreta di status gerarchico superiore richiede l'uso di strutture che si trovano nella parte anteriore delle cortecce temporali, e l'accesso a una conoscenza di livello gerarchico inferiore implica strutture che si trovano nelle cortecce occipitali posteriori. Questo requisito necessario risiede nelle connessioni e nelle loro interazioni, e non implica necessariamente una localizzazione precisa e puntuale in un tessuto nervoso specifico (Damasio, 1994).
L'ipotesi di una distinzione tra regioni del cervello topografiche e amodali non richiede necessariamente caratteristiche anatomiche differenziate. In altre parole, esistono svariate caratteristiche anatomiche che sembrano essere correlate con questa ipotesi e sono, per esempio, la natura distintiva delle cortecce primarie (che sono le uniche a ricevere cospicue connessioni talamo-corticali e ad avere una forte mappatura topografica), la natura distribuita delle connessioni, la fitta rete di connessioni dirette e indirette, che possono agevolmente codificare una varietà di operazioni concettuali altrettanto complessa e, infine, l'esistenza di interazioni temporali sincrone (v. oltre).
Selezione di gruppi neuronali
Un nuovo aspetto dell'approccio basato sulle zone di convergenza sta nel considerare le combinazioni di connessioni come codici di legame che servono per le relazioni cognitive dinamiche. Diverse altre proposte di organizzazioni su larga scala hanno affrontato in modi differenti il problema della natura dei codici neuronali e delle rappresentazioni. La teoria della selezione neuronale sviluppata da G. Edelman (Edelman, 1987; 1993; Sporns e Tononi, 1994) propone che la formazione, l'adattamento e le interazioni fra collezioni locali di gruppi neuronali siano regolate da meccanismi di selezione, piuttosto che da meccanismi di istruzione. La correlazione o la congiunzione di gruppi neuronali viene vista come un processo dinamico, portato avanti da connessioni rientranti. Il concetto di connessione 'rientrante' implica l'esistenza di reti con connessioni reciproche feedforward e ricorrenti, ma il termine è anche compatibile con le connessioni collaterali orizzontali intrinseche e con le reti di connessioni prese come aggregati (fig. 25). Una componente chiave è semplicemente la densità del plesso di connessioni, in cui si può realizzare un numero enorme di combinazioni neurali attraverso un'organizzazione parallela nello spazio e nel tempo. Le interazioni rientranti sono state di recente applicate in un modello per la visione del colore (Wray ed Edelman, 1996).
Gruppi neuronali correlati temporalmente
Le codifiche di popolazione o di gruppo, sebbene forniscano delle soluzioni possibili per le rappresentazioni di un numero elevato di caratteristiche degli stimoli, pongono però il problema di come la rappresentazione di un certo contenuto possa essere identificata in modo non ambiguo come appartenente a un insieme (Singer, 1994; Sporns e Tononi, 1994; Singer, 1995a). Un meccanismo biologicamente plausibile, che ha ricevuto un notevole sostegno dai dati sperimentali, è quello basato sulla correlazione temporale dell'attività di gruppi di neuroni distribuiti nello spazio (Singer, 1993). È stata dimostrata la presenza di sincronizzazione temporale tramite l'analisi dei diagrammi di correlazione incrociata, ottenuta mediante registrazioni con elettrodi multipli. Una recente interpretazione di queste scoperte suggerisce che la corteccia adotti due strategie complementari: da un lato, l'analisi delle relazioni frequenti e rilevanti dal punto di vista del comportamento, condotta da gruppi di cellule con proprietà fisse di risposta non molto selettive ('precablate'); dall'altro, l'associazione dinamica di queste cellule in gruppi funzionalmente coerenti. Si pensa che dietro a queste due operazioni vi siano rispettivamente le connessioni corticali feedforward e le connessioni intrinseche assocIatIve, ovvero le connessioni laterali locali dei neuroni piramidali (Singer, 1995b). La sincronizzazione temporale fornisce un altro quadro specifico per le teorie sull' organizzazione generale del cervello. Poiché essa è una componente largamente prevalente delle funzioni neuronali, la sincronizzazione può anche essere inserita in altri contesti, come la selezione neuronale e l'architettura delle zone di convergenza.
Conclusioni
In questo saggio sono state discusse alcune delle componenti della struttura del cervello e il modo in cui queste potrebbero interagire, in architetture su larga scala, come substrati di funzioni cognitive. La relazione tra la struttura e la funzione neuronale, sebbene indubbiamente ordinata, non è necessariamente chiara o diretta, esattamente come raramente esiste un singolo gene per una malattia o un singolo gene che produce un comportamento (come il 'gene della violenza'). Si pensi anche al sistema motorio, dove gli stessi muscoli o gruppi di muscoli possono generare una grande varietà di movimenti. Inoltre, gli studi di visualizzazione funzionale hanno suggerito che vi sia un certo grado di flessibilità nelle potenzialità funzionali del cervello. Per esempio, la corteccia visiva primaria è coinvolta in altri processi oltre a quelli che riguardano la percezione, e potrebbe giocare un ruolo importante nei processi di immaginazione o di rievocazione di informazioni visive. Una sfida importante per il futuro sarà quella di capire meglio queste complesse relazioni e di sviluppare ulteriormente un linguaggio appropriato per esprimerle.
Bibliografia citata
AMIT, D.J., BRUNEL, N. (1997) Model of global spontaneous activity and local structured activity during delay periods in cerebral cortex. Cereb. Cortex, 7, 237-252.
BLASDEL, G.G. (1992) Differential imaging of ocular dominance and orientation selectivity in monkey striate cortex. J. Neurosci., 12, 3115-3138.
BLASDEL, G.G., LUND, J.S. (1983) Termination of afferent axons in macaque striate cortex. J. Neurosci., 3, 1389-1413.
BRITTEN, K.H., SHADLEN, M.N., NEWSOME, W.T., MOVSHON, J.A. (1992) The analysis of visual motion: a comparison of neuronal and psychophysical performance. J. Neurosci., 12, 4745-4765.
BULLIER, J., GIRARD, P., SALIN, P.A. (1994) The role of area 17 in the transfer of information to extrastriate visual cortex. In Cerebral cortex, a c. di Jones E.G. e Peters A., vol. 10, New York, Plenum Press, pp. 301-330.
BULLIER, J., HUPÉ, J.M., JAMES, A.C., GIRARD, P. (1996) Functional interactions between areas VI and V2 in the monkey. J. Physiol., 90, 217-220.
DAMASIO, A.R. (1989) The brain binds entities and events by multiregional activation from convergence zones. Neural Comput., l, 123-132.
DAMASIO, A.R. (1994) Descartes' error: emotion, reason, and the human brain. New Y ork, G.P. Putnam.
DAMASIO, A.R., DAMASIO, H., VAN HOESEN, G.W. (1982) Prosopagnosia: anatomie basis and behavioral mechanisms. Neurology, 32, 331-341.
DAMASIO A.R., DAMASIO, H. (1994) Cortical systems for retrieval of concrete knowledge: the convergence zone framework. In Large-scale neuronal theories of the brain, a c. di Koch Ch., Davis J.L., Cambridge, Mass., MIT Press, pp. 61-74.
DEFELIPE, J., FARINAS, I. (1992) The pyramidal neuron of the cerebral cortex: morphological and chemical characteristics of the synaptic inputs. Prog. Neurobiol., 39, 563-607.
DOUGLAS, R., MARTIN, K. (1998) Neocortex. In The synaptic organization of the brain, a c. di Shepherd G.M., Oxford-New York, Oxford University Press, pp. 459-509.
EDELMAN, G.M. (1987) Neural darwinism: the theory of neuronal group selection. New York, Basic Books.
EDELMAN, G.M. (1993) Neural darwinism: selection and reentrant signaling in higher brain function. Neuron, 10, 115-125.
FELLEMAN, D.J. VAN ESSE N, D.C. (1991) Distributed hierarchical processing in the primate cerebral cortex. Cereb. Cortex, l, 1-47.
GILBERT, C.D. (1992) Horizontal integration and cortical dynamics. Neuron, 9, 1-13.
GRINVALD, A., LIEKE, E.E., FROSTIG, R.D., HILDESHEIM, R. (1994) Cortical point-spread function and long-range lateral interactions revealed by real-time optical imaging of macaque monkey primary visual cortex. J. Neurosci., 14, 2545-2568.
HIGUCHI, S., MIYASHITA, Y. (1996) Formation of mnemonic neuronal responses to visual paired associates in inferotemporal cortex is impaired by perirhinal and entorhinal lesions. Proc. Natl. Acad. Sci. USA, 93, 739-743.
HIRSCH, J.A., GILBERT, C.D. (1991) Synaptic physiology ofhorizontal connections in the cat's visual cortex. J. Neurosci., Il, 1800-1809.
HOFSTADTER, D.R. (1984) GodeI, Escher, Bach: un'eterna ghirlanda brillante. Milano, Adelphi.
HUBEL, D.H., WIESEL, T.N. (1977) Ferrier lecture. Functional architecture of macaque monkey visual cortex. Proc. R. Soc. Lond. B Biol. Sci., 198, l-59.
IIJIMA, T., WITTER, M.P., ICHIKAWA, M., TOMINAGA, T., KAJIWARA, R., MATSUMOTO, G., (1996) Entorhinal-hippocampal interactions revealed by real-time imaging. Science, 272, 1176-1179.
KLEINFELD, D., DELANEY, K.R. (1996) Distributed representation of vibrissa movement in the upper layer of somatosensory cortex revealed with voltage-sensitive dyes. J. Comp. Neurol., 375, 89-108.
KOCH, CH., DAVIS, J.L., a C. di (1994) Large-scale neuronal theories of the brain. Cambridge, Mass., MIT Press.
LEVITT, J.B., BARBE, M.F., EAGLESON, K.L. (1997) Patteming and specification of the cerebral cortex. Annu. Rev. Neurosci., 20, 1-24.
MUMFORD, D. (1992) On the computational architecture of the neocortex. II. The role of cortico-corticalloops. Biol. Cybern., 66, 241-251.
NICOLELIS, M.A.L., BACCALA, L.A., LIN, R.C., CHAPIN, J.K. (1995) Sensorimotor encoding by synchronous neural ensemble activity at multiple levels of the somatosensory system. Science, 268, 1353-1358.
NOWAK, L.G., BULLIER, J. (1997) The timing ofinformation transfer in the visual system. In Cerebral cortex, a c. di Jones E.G. e Peters A., vol.12, New York, Plenum Press, pp. 205-242.
PETERS, A., SETHARES, C. (1991) Organization of pyramidal neurons in area 17 of monkey visual cortex. J. Comp. Neurol., 306, 1-23.
ROCKLAND, K.S. (1997) Elements of cortical architecture: hierarchy revisited. In Cerebral cortex, a c. di Jones E.G. e Peters A., vol. 12, New York, Plenum Press, pp. 243-294.
SALIN, P.A., BULLIER, J. (1995) Cortico-cortical connections in the visual system: structure andfunction.Physiol. Rev., 75, 107-154.
SALZMAN, C.D., MURASUGI, C.M., BRITTEN, K.H., NEWSOME, W.T. (1992) Microstimulation in visual area MT: effects on direction discrimination performance. J. Neurosci., 12, 2331-2355.
SINGER, W. (1993) Synchronization of cortical activity and its putative role in information processing and leaming. Annu. Rev. Physiol., 55, 349-374.
SINGER, W. (1994) Putativefunctions of the temporal correlations in neocortical processing. In Large-scale neuronal theories of the brain, a c. di Koch Ch., Davis J.L., Cambridge, Mass., MIT Press, pp. 201-237.
SINGER, W. (1995a) Time as coding space in neurocortical processing: a hypothesis. In The cognitive neurosciences, a c. di Gazzaniga M.S., Cambridge, Mass., MIT Press, pp. 91-104.
SINGER, W. (1995b) Development and plasticity of cortical processing architectures. Science, 270, 758-764.
SPORNS, O., TONONI, G. (1994) Selectionism and the brain, San Diego, Academic Press.
TANAKA, K. (1997) Columnar organization in the inferotemporal cortex. In Cerebral cortex, a c. di Jones E.G. e Peters A., vol. 12, New York, Plenum Press, pp. 469-498.
ULLMAN, S. (1995) Sequence seeking and counter streams: a computational model for bidirectional information flow in the visual cortex. Cereb. Cortex, 5, 1-11.
WELKER, E., ARMSTRONG-JAMES, M., BRONCHTI, G., OUREDNIK, W., GHEORGHITA-BAECHLER, F., DUBOIS, R., GUERNSEY, D.L., VAN DER LOOS, H., NEUMANN, P.E. (1996) Altered sensory processing in the somatosensory cortex of the mouse mutant barrelless. Science, 271, 1864-1867.
WHITE, E.L. (1989) Cortical circuits: synaptic organization of the cerebral cortex-structure, Junction, and theory. Boston, Birkhiiuser.
WILSON, M.A., TONEGAWA, S. (1997) Synaptic plasticity, pIace cells and spatial memory: study with second generation knockouts. Trends Neurosci., 20, 102-106.
WOOLSEY, T.A., VAN DER LOOS, H. (1970) The structuralorganization of layer IV in the somatosensory region (SI) of mouse cerebral cortex: description of a cortical field composed of discrete cytoarchitectonic units. Brain Res., 17, 205-242.
WRAY, J., EDELMAN, G.M. (1996) A model of color vision based on cortical reentry. Cereb. Cortex, 6, 701-716.
Bibliografia generale
BRAITENBERG, V., SCHUZ, A. Anatomy of the cortex. Statistics and geometry. New York, Springer Verlag, 1991.
DAMASIO, A.R. Descartes' error: emotion, reason, and the human brain. New York, G.P. Putnam, 1994.
EDELMAN, G.M. Neural darwinism: the theory of neuronal group selection. New York, Basic Books, 1987.
FRACKOWIAK, R.S.J., FRISTON, K.J., FRITH, C.D., DOLAN, R.J., MAZZIOTTA, J.C. Human brain function. NewYork, Academic Press, 1997.
GAZZANIGA, M.S., a c. di, The cognitive neurosciences. Cambridge, Mass., MIT Press, 1995.
JONES, E.G., PETERS A., a c. di, Cerebral cortex. 12 voll. New Y ork, Plenum Press, 1984-1997.
WHITE, E.L. Cortical circuits: synaptic organization of the cerebral cortex-structure, Junction, and theory. Boston, BirkMuser, 1989.
ZEKI, S. A vision of the brain. Londra, Blackwell Scientific Publications, 1993.
© Istituto della Enciclopedia Italiana - Riproduzione riservata


